2020.05.04.25
Files > Volume 5 > Vol 5 No 4 2020
Mechanisms of Nuclear Transport in the cell: RNA exosome in Saccharomyces cerevisiae

Bruna Rech1 and Fernando A. Gonzales-Zubiate2
Available from: http://dx.doi.org/10.21931/RB/2020.05.04.25
ABSTRACT
Ribonucleases (RNases) functions in the cell include precise maturation of non- coding RNAs and degradation of specific RNA transcripts that are no longer necessary. RNAses are present in the cell as single units or assembled as multimeric complexes; one of these complexes is the RNA exosome, a highly conserved complex essential for RNA processing and degradation. In the yeast Saccharomyces cerevisiae, the RNA exosome comprises eleven subunits, two with catalytic activity: Rrp6 and Rrp44, where the Rrp6 subunit is exclusively nuclear. Despite the RNA exosome has been intensively investigated since its discovery in 1997, only a few studies were accomplished concerning its nuclear transport. This review describes recent research about cellular localization and transport of this essential complex.
Keywords: Ribonucleases, exosome, nuclear transport, Rrp6, Rrp44, RNA processing, RNA degradation.
INTRODUCTION
RNA exosome is a multiprotein RNase involved in the modification of all RNAs in the cell; its primary function includes exonucleolytic and endonucleolytic cleavage of pre-ribosomal RNA (pre-rRNA), messenger RNA (mRNA), small nuclear RNA (snRNA), and small nucleolar RNA (snoRNA)1. Other RNA exosome substrates were described and included a significant group of long noncoding RNAs (lncRNAs): cryptic unstable transcripts (CUT) in yeast and upstream promoter transcripts (PROMPT) in humans, prematurely terminated RNA (ptRNA), enhancer RNA (eRNA) and long intergenic RNA (lincRNA)2 (Figure 1). Highly conserved throughout evolution, exosome complex can be found in several species such as Archaea, Saccharomyces cerevisiae, Drosophila melanogaster, Arabidopsis thaliana, Mus musculus, and humans, to mention some. In Saccharomyces cerevisiae, the exosome is present in both nucleus and cytoplasm, fulfilling different functions depending on its substrates. Two structures compose the exosome core: a barrel-like with six different subunits, each containing an inactive RNase PH domain (Rrp41, Rrp42, Rrp43, Rrp45, Rrp46, Mtr3); and a cap, capping this barrel with three RNA binding subunits (Rrp4, Rrp40, Csl4). The catalytic subunits bind to opposite sides of the core: Rrp44 with endo- and exonucleolytic activities is present in both nuclear and cytoplasmic exosome3-5; Rrp6 with a 3'-5' exonucleolytic activity present only in the nuclear exosome6 (Figure 2).
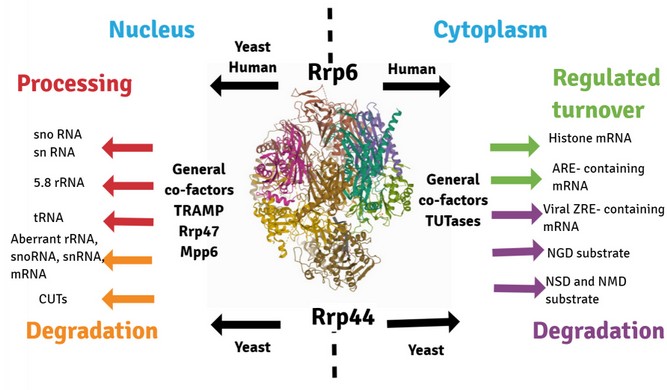
Figure 1. RNA quality control by the exosome for aberrant mRNA, tRNA, rRNA, and other noncoding RNA (ncRNA) species in every cell compartment. Arrows indicate the localization of active nucleases Rrp6 and Rrp44. TRAMP (complex consists of helicase Mtr4, polyA polymerase Trf4 or Trf5, and RNA-binding proteins Air1 or Air2), TUTases (terminal RNA uridylyltransferases). CUTs (Cryptic unstable transcripts), upstream promoter transcripts (PROMPTs), and upstream noncoding transcripts (UNTs). ARE (AU-rich instability element). Aberrant mRNAs with translation defects: NMD (Nonsense-mediated decay), NSD (non-stop decay), and NGD (no-go decay).
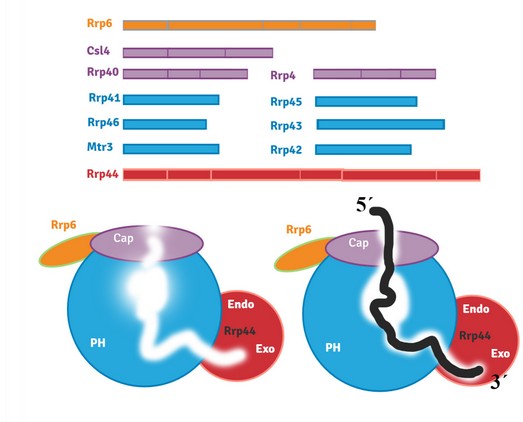
Figure 2. Exosome structure in yeast. A. Cap and barrel-like structures of exosome in purple and blue, respectively. Associated nucleases are in red (Rrp44) and orange (Rrp6). B. RNA-free form of the exosome. C. RNA going through the exosome channel to be processed.revistabionatura.com/2020.05.04.25.html
The nuclear exosome7,8 processes degradation of incorrectly processed RNAs and maturation of ribosomal RNAs, small nuclear RNAs and small nucleolar RNAs. Furthermore, cytoplasmic exosome plays a vital role in regulating mRNA biogenesis's gene expression by mRNA degradation9,10.
Nuclear transport of exosome
Nuclear transport is primarily mediated by specific interactions between a karyopherin, a protein transporter, and signal sequences (NLS; Nuclear Localization Signal) present in the cargo proteins 11,12 (Figure 3). In Saccharomyces cerevisiae, there is one karyopherin: Srp1. Besides, there are 14 karyopherins β: ten of which are involved in transport to the nucleus (importins - Kap95, Kap104, Sxm1/Kap108, Mtr10, Kap114, Nmd5, Kap120/Lph2, Pse1/Kap121, Kap122, Kap123), three are involved in transport to the cytoplasm (exportins - Cse1, Crm1/Xpo1, Los1), and one karyopherin is involved in transport in both directions, Msn513. Interaction of karyopherins Srp1 and Kap95 with Rrp6 were observed initially in protein purification experiments 14-17 and co-immunoprecipitation experiments using Rrp6 as bait showing a surprising number of karyopherins: Kap95, Srp1, Kap114, Kap123, Sxm1, and Cse118.
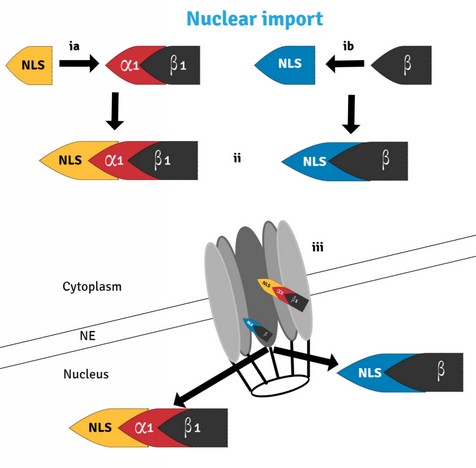
Figure 3. Nuclear transport through the Nuclear Pore Complex requires the recognition and binding of NLS-containing cargo proteins by either the importins α/β1 heterodimer (ia) or importin β alone (ib) on the cytoplasm. Once bound (ii), the transport complex moves through the NPC (iii) via a series of transient interactions between importin β and nucleoporins in the NPC
Presence of NLSs in exosome subunits
In-silico and experimental analysis showed three subunits containing NLSs in their sequence: Rrp6, Rrp43, and Rrp44. Subunit Rrp43 contains a potential NLS between the residues 203 and 213: 203LKMKRKWSYVL213 identified using the software NLS Mapper and NLStradamus 19,20. No experiments are available to test if this motif could serve as a nuclear localization signal.
Subunit Rrp6 showed the presence of a three different NLSs in its sequence:
NLS1 sequence 104NSKSRGSDLQYLGEFSGKNFSPTKRVEKP132 identified by NLS Mapper and NLStradamus 19,20. NLS2: 153KEKPNALKPLSESLRLVDDDENNPSHYPHPY183, contains a multipartite proline–tyrosine nuclear localization signal called PY-NLS with a C- terminal R/H/K-X2–5-P-Y motif within a positively charged region of approximately 30 amino acids 18,21. They could be recognized by importins Kap104, Sxm1/Kap108, Kap121, Kap114, Nmd5/Kap119, and Kap95 22-24.
NLS3: 697RQQKKRRFDPSSSDSNGPRAAKKRRPA723, a classical NLS at the C-terminal region of Rrp6 that could be recognized by importin α Srp125. It was observed that lack of any of the mentioned NLSs in the Rrp6 sequence lead to a partial mislocalization in the cell18.
Subunit Rrp44 showed to possess three NLSs
NLS1: 172RAIRKTCQWYSEHLKPY188, similar to Rrp6-NLS2 with a nuclear localization signal PY-NLS, NLS1 is located PinC domain of Rrp4426.
NLS2 is located in the cold shock domain 2 CSD226 between the residues 370 and 401 of Rrp44 370RRLLAKDAMIAQRSKKIQPTAKVVYIQRRSWR401 and contains the consensus NLS R/K-X2-L-XnV/Y-X2-V/I-X-K/R-X3-K/R that can be recognized by the importins Kap114, Kap95, Kap123, Pse1, and Kap10427.
NLS3 is located at the C-terminus of Rrp44: 988DPITSKRKAELLLK1001. This NLS is similar to the NLS in Drosophila Rrp44 (dDis3), which is recognized by importin-3 28,29. From the three NLSs presents in Rrp44 sequence, NLS1 is the most critical signal for Rrp44 nuclear import. NLS1, together with either NLS2 or NLS3 is sufficient for Rrp44 nuclear localization18.
The presence of three nuclear localization signals in both Rrp6 and Rrp44 and more of one karyopherin involved in their nuclear transport confirm overlapping and redundancy in the import pathways.
Depletion of karyopherins affect the localization of exosome subunits
Depletion of essential karyopherin Srp1 in the cell after 14 hours showed Rrp6 visualized in the cytoplasm, although Rrp6 remained concentrated in the nucleus; levels of Rrp6 decreased upon inhibition of Srp1 expression, it could suggest when not efficiently transported to the nucleus, Rrp6 may be destabilized18. Inhibition of karyopherin Kap95 expression for 14 hours strongly affected Rrp6 localization; Rrp6 was observed in the cytoplasm and concentrated in the nucleus. Surprisingly, the deletion of the karyopherin Sxm1/Kap108 gene affected the localization of Rrp6 partially Sxm1 is not essential for growth but Rrp6 was detected in the cytoplasm, and its mislocalization was more robust at 37°C. These results showed that Rrp6 could associate with different importins for its transport to the nucleus18.
Localization of Rrp44 in wild-type cells is mainly in the nucleus with a weak signal in the cytoplasm. The depletion of Srp1 or Kap95 levels in the cell leads to Rrp44 partial mislocalization, evidenced by a strong signal in the cytoplasm. Unlike Rrp6, the depletion of Sxm1 did not show any effect on Rrp44 localization30.
CONCLUSION
The RNA exosome is localized in the cytoplasm and the nucleus (specifically in the nucleolus). Although its predominantly nuclear localization, only three exosome subunits contain NLSs in their sequence, and from these three, two have shown to be transported by karyopherins: Rrp6 and Rrp44. It is already known that Srp1 and Kap95 are the karyopherins involved in the nuclear transport of Rrp44 and Rrp6, with the extra help of karyopherin Sxm1 for Rrp6 transport. However, it is uncertain how the rest of the subunits are transported to the nucleus or transported independently or as a complex. Only further studies will help to elucidate these questions and better understand the transport mechanisms of this complex.
REFERENCES
1. Allmang C, Kufel J, Chanfreau G, Mitchell P, Petfalski E, Tollervey D. Functions of the exosome in rRNA, snoRNA and snRNA synthesis. EMBO J [Internet]. 1999 Oct 1;18(19):5399–410. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1171609&tool=pmcentrez&rendertype=abstract
2. Ogami K, Chen Y, Manley JL. RNA surveillance by the nuclear RNA exosome: Mechanisms and significance. Noncoding RNA. 2018;4(1).
3. Liu Q, Greimann JC, Lima CD. Reconstitution, activities, and structure of the eukaryotic RNA exosome. Cell [Internet]. 2006 Dec 15 [cited 2012 Apr
8];127(6):1223–37. Available from:http://www.ncbi.nlm.nih.gov/pubmed/17174896
4. Makino DL, Baumgärtner M, Conti E. Crystal structure of an RNA- bound 11-subunit eukaryotic exosome complex. Nature [Internet]. 2013 Mar 7 [cited 2013 Nov 5];495(7439):70–5. Available from: http://www.ncbi.nlm.nih.gov/pubmed/23376952
5. Dziembowski A, Lorentzen E, Conti E, Séraphin B. A single subunit, Dis3, is essentially responsible for yeast exosome core activity. Nat Struct Mol Biol. 2007;14(1):15–22.
6. Burkard KTD, Butler JS. A Nuclear 3 ′ -5 ′ Exonuclease Involved in mRNA Degradation Interacts with Poly ( A ) Polymerase and the hnRNA Protein Npl3p A Nuclear 3Ј-5Ј Exonuclease Involved in mRNA Degradation Interacts with Poly ( A ) Polymerase and the hnRNA Protein Npl3p. 2000;20(2).
7. Houseley J, LaCava J, Tollervey D. RNA-quality control by the exosome. Nat Rev Mol Cell Biol [Internet]. 2006;7(7):529–39. Available from: http://www.ncbi.nlm.nih.gov/pubmed/16829983
8. Allmang C, Kufel J, Chanfreau G, Mitchell P, Petfalski E, Tollervey D. Functions of the exosome in rRNA, snoRNA and snRNA synthesis. EMBO J [Internet]. 1999 Oct 1;18(19):5399–410. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi? artid=1171609&tool=pmcentrez&rendertype=abstract
9. Anderson JSJ, Parker R. The 3´ to 5´ degradation of yeast mRNAs is a general mechanism for mRNA turnover that requires the SKI2 DEVH box protein and 3´ to 5´ exonucleases of the exosome complex. 1998;17(5):1497–506.
10. Oliveira CC, Gonzales FA, Zanchin NIT. Temperature-sensitive mutants of the exosome subunit Rrp43p show a deficiency in mRNA degradation and no longer interact with the exosome. Nucleic Acids Res [Internet]. 2002 Oct 1;30(19):4186–98. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi? artid=140545&tool=pmcentrez&rendertype=abstract
11. Xu D, Farmer A, Chook YM. Recognition of nuclear targeting signals byKaryopherin-β proteins. Curr Opin Struct Biol. 2010;20(6):782–90.
12. Lange A, Mills RE, Lange CJ, Stewart M, Devine SE, Corbett AH. Classical nuclear localization signals: Definition, function, and interaction with importin α. J Biol Chem. 2007;282(8):5101–5.
13. Mosammaparast N, Pemberton LF. Karyopherins: from nuclear-transport mediators to nuclear-function regulators. Trends Cell Biol [Internet]. 2004 Oct [cited 2013 Nov 5];14(10):547–56. Available from: http://www.ncbi.nlm.nih.gov/pubmed/15450977
14. Ho Y, Gruhler A, Heilbut A, Bader GD, Moore L, Adams S, et al. Systematic identification of protein complexes in Saccharomyces cerevisiae by mass spectrometry. 2002;415(January):2–5.
15. Gavin A-C, Bösche M, Krause R, Grandi P, Marzioch M, Bauer A, et al.Functional organization of the yeast proteome by systematic analysis of protein complexes. Nature [Internet]. 2002 Jan 10;415(6868):141–7. Available from:
http://www.ncbi.nlm.nih.gov/pubmed/11805826
16. Gavin A-C, Aloy P, Grandi P, Krause R, Boesche M, Marzioch M, et al. Proteome survey reveals modularity of the yeast cell machinery. Nature [Internet]. 2006 Mar 30 [cited 2014 Jan 29];440(7084):631–6. Available from: http://www.ncbi.nlm.nih.gov/pubmed/16429126
17. Synowsky S a, van Wijk M, Raijmakers R, Heck AJR. Comparative multiplexed mass spectrometric analyses of endogenously expressed yeast nuclear and cytoplasmic exosomes. J Mol Biol [Internet]. 2009 Jan 30 [cited 2013 Nov 5];385(4):1300–13. Available from:http://www.ncbi.nlm.nih.gov/pubmed/19046973
18. Gonzales-Zubiate FA, Okuda EK, Da Cunha JPC, Oliveira CC. Identification of karyopherins involved in the nuclear import of RNA exosome subunit Rrp6 in Saccharomyces cerevisiae. J Biol Chem. 2017;292(29):12267–84.
19. Kosugi S, Hasebe M, Tomita M, Yanagawa H. Systematic identification of cell cycle-dependent yeast nucleocytoplasmic shuttling proteins by prediction of composite motifs. Proc Natl Acad Sci U S A. 2009;106(14):10171–6.
20. Nguyen Ba AN, Pogoutse A, Provart N, Moses AM. NLStradamus: a simple Hidden Markov Model for nuclear localization signal prediction. BMC Bioinformatics. 2009;10:202.
21. Lee BJ, Cansizoglu AE, Suel KE, Louis TH, Zhang Z, Chook YM. Rules for Nuclear Localization Sequence Recognition by Karyopherin b2. Cell. 2006;126(3):543–58.
22. Suel KE, Gu H, Chook YM. Modular Organization and Combinatorial Energetics of Proline – Tyrosine Nuclear Localization Signals. PLoS Biol. 2008;6(6).
23. Lee C, Hodgins DC, Calvert JG, Welch SKW, Jolie R, Yoo D. The nuclear localization signal of the PRRS virus nucleocapsid protein modulates viral replication in vitro and antibody response in vivo. Adv Exp Med Biol. 2006;581:145–8.
24. Süel KE, Chook YM. Kap104p imports the PY-NLS-containing transcription factor Tfg2p into the nucleus. J Biol Chem. 2009;284(23):15416–24.
25. Phillips S, Butler JS. Contribution of domain structure to the RNA 3 end processing and degradation functions of the nuclear exosome subunit Rrp6p.
2003;1098–107.
26. Lebreton A, Tomecki R, Dziembowski A, Séraphin B. Endonucleolytic RNA cleavage by a eukaryotic exosome. Nature [Internet]. 2008 Dec 18 [cited 2012 Mar 11];456(7224):993–6. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19060886
27. Fries T, Betz C, Sohn K, Caesar S, Schlenstedt G, Bailer SM. A novel conserved nuclear localization signal is recognized by a group of yeast importins. J Biol Chem. 2007;282:19292–301.
28. Graham AC, Davis SM, Andrulis ED. Interdependent nucleocytoplasmic trafficking and interactions of Dis3 with Rrp6, the core exosome, and importin- α3. 2009;10(5):368–85.
29. Mamolen M, Smith A, Andrulis ED. Drosophila melanogaster Dis3 N- terminal domains are required for ribonuclease activities, nuclear localization and exosome interactions. Nucleic Acids Res. 2010;38(16):5507–17.
30. Okuda EK, Gonzales-Zubiate FA, Gadal O, Carla C. Oliveira. Nucleolar localization of the yeast RNA exosome subunit Rrp44 hints at early pre-rRNA processing as its main function. J Biol Chem. 2020;(1):1–9.
Received: 10 August 2020
Accepted: 16 October 2020
Bruna Rech1 and Fernando A. Gonzales-Zubiate2
1Fertility Medical Group, Sao Paulo, Brazil. https://orcid.org/0000-0002-2690-5057
2School of Biological Sciences and Engineering, Yachay Tech University, Ecuador.
https://orcid.org/0000-0001-9519-4079