2021.06.02.25
Files > Volume 6 > Vol 6 No 4 2021 > Vol 6 No 2 2021
INVESTIGATION / RESEARCH
Effect of Plasmodium berghei infection on fetuses in pregnant BALB/c mice at two periods of pregnancy

Andreina Gómez2, Beatriz Pernía3, Lizbeth Zamora1 and Lilian M. Spencer *1,2
Available from: http://dx.doi.org/10.21931/RB/2021.06.02.25
ABSTRACT
Malaria is a disease caused by a protozoan of the genus Plasmodium in humans and vertebrates. It has a high morbidity and mortality rate, especially in pregnant women living in countries with high transmission rates. Murine models have been an excellent tool to evaluate the effects of malarial infection in the mother-fetus relationship. For this reason, we evaluated the effect of malarial infection on fetal development at the beginning and middle of the gestational period in BALB/c mice infected with Plasmodium berghei ANKA. Our results show that malarial infection at the beginning of pregnancy markedly affects the development of the fetus in size, weight, and development of its limbs so that the control of the pregnant mother is relevant at the beginning of gestation
Keywords: malaria, loss weight, rodent, gestational period
INTRODUCTION
Malaria is an endemic tropical disease with a high incidence among women and children. It is therefore considered a significant public health problem. World Health Organization (WHO) reported pregnant women in sub-Saharan Africa to show high malarial infection1. In 2019, WHO estimates 12 million pregnancies with malaria infection and 822.000 children with low birth weight2. Reflecting a high morbidity and mortality rate after infection as medical complications such as anemia, intrauterine growth retardation, low birth weight (LBW), and neonatal death occur 3,4. The vulnerability of pregnant women to malaria increases in the first and second trimesters of the gestation period. However, there is some debate as to whether hormonal and immunological changes increase malaria sensitivity in pregnancy5. As a result, prenatal diagnosis and control are complex. Besides, known drugs are less effective due to resistance generated by parasites 6.
Malaria infection in pregnant women occurs when intracellular Plasmodium falciparum parasites accumulate and interact with chondroitin sulfate A (CSA) receptors, resulting in placental malaria5,6. The malaria-infected placenta has a thickened trophoblastic basement membrane that acts as a protective measure for the baby against increased cytokines that damage the placental tissue. Therefore, morphological alterations of the membrane directly affect the pathological conditions of the fetus7. The placenta has parasites that express antigens on its surface, known as variant surface antigens (VSA)5. The VSA of the malaria parasites prevents them from being recognized by the human immune system as they belong to the Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1)8.
Meanwhile, the VAR2CSA protein present in malaria-infected red blood cells recognizes CSA in pregnant women, causing it to accumulate in the placenta. The parasites infect other red blood cells that will accumulate in the placenta9. Chronic malaria infection is related to low birth weight by restricting space for development, while the acute infection is related to early termination of pregnancy10.
Plasmodium falciparum infection is the type of malaria with the highest incidence in pregnant women because infected red blood cells bind more quickly to the placenta during the gestation period11,12. Several experimental models of malaria infection reproduce the immune responses of the disease. Rats and mice are infected with Plasmodium vinckei, Plasmodium yoelii and Plasmodium berghei, among others13. To study malaria in pregnant women, the Plasmodium berghei ANKA strain is used in murine animals because it mimics the infectious process of chronic malaria in the gestation stage that occurs with Plasmodium falciparum in humans 10. Specifically, pregnant BALB/c mice infected with Plasmodium berghei develop complications such as decreased fetal size, premature birth, slowed uterine growth, and postnatal growth problems 13,14. The factors presented above are related to the placenta's nutritional deficit, which is essential for a healthy pregnancy 15, 16.
This work aimed to determine in which gestation period the malarial infection is more deleterious to fetal development at the beginning or middle of gestation. For this, we evaluated the weight, size, and limb development in fetuses from pregnant BALB/c female mice.
MATERIALS AND METHOD
Experimental infection and appearance of the female BALB/c mice
Mice reared under specific pathogen-free conditions and with food and water ad libitum in the animal laboratory at the Simón Bolívar University. Female mice of 12-week-old were mate with a young puberty male mouse. When the formation of the vaginal plug was verified the next day, the male was separated from females in each group17. In this study, pregnant females were divided into three experimental groups with six mice per group (Figure 1 presents the experiments with mice). One control group that was not infected with Plasmodium berghei ANKA strain and two groups that were infected with the parasite at the beginning (second day after of formation vaginal plug) of gestational and in the middle of the gestational period (eighth day after of formation vaginal plug).

Figure 1. Experimental mice groups to evaluate the parasitaemia and physical conditions of fetuses. One control group was not infected with the ANKA strain of Plasmodium berghei, and two groups infected with the parasite at the beginning (second day) of gestational and in the middle of the gestational period (eighth day).
The parasite used for the challenge was the lethal ANKA strain of P. berghei; the parasite was stored at -80°C and passaged once in a mouse before use in these experiments. When the parasitaemia was 40%, parasitized red blood cells (PRBCs) were isolated using a cellulose column (CF-11, Sigma, with NaCl 0.85% solution). After, the trophozoites and schizonts were isolated by a discontinuous Percoll column (60, 70, and 80% gradient in ascending order)18.
The parasite challenge was administered by intravenous injection of five thousand PRBCs to two experimental groups at the 2nd and other at the 8th days of the gestational period. Six mice were placed for each group and the control group that were not pregnant but if infected with P. berghei. Erythrocytic stage parasitaemia was assessed daily on smears made from tail blood from infected mice and stained with Giemsa's reagent to determine percentage of parasitaemia. These experiments were carried out twice to evaluate the effect of parasite infection on the fetuses.
In summary, we conducted two experimental groups of pregnant and infected mice. One of them to determine parasitemia and the other to determine the conditions of the fetuses depending on the day of infection with the parasite
Fetus extraction and evaluation in size and weight
At 7 days post-infection, pregnant mice of the groups infected with P. berghei on the 2nd and 8th day of gestation were sacrificed, and their uterine sacs were opened (using a longitudinal incision on the mesometrial side); each mouse pregnant had an average of 10±1 fetus. Then, we macroscopically evaluated the fetuses. Which were washed with a physiological solution and weighed to determine their weight on balance Sartorius Entris II model (g), fetal length with a calibrate Vernier (cm), and external anatomical characteristics such as the development of the extremities.
The experiments described in this study were carried out following the laboratory's standards and the Bioethics Committee of the Simón Bolívar University.
Statistical analysis
To analyze whether the data presented a normal distribution, both Anderson-Darling and Levene tests for homoscedasticity were applied. Subsequently, to compare the percentage of parasitaemia between treatments, a two-way ANOVA test (p <0.001) was applied, followed by a Dunnet test. Then, to compare the fetuses' weight and length, a non-parametric Kruskall-Wallis test was applied (p <0.001). The graphics were made using the R program version 4.0.2.
RESULTS
Figure 2 shows the percentage of parasitaemia during the infection, where it is evident that pregnant mice presented higher percentages of parasitaemia than non-pregnant control mice. On day 7 after infection, a significant difference is clearly shown between the three groups of mice infected with the parasite (F = 46.28; p ≤ 0.001). With a percentage of parasitaemia of 60.0 ± 10.0% (green line) in mice inoculated on day 8 and 33.3 ± 5.8% in those inoculated on the second day (pink line), compared to control 6.8 ± 2.0% (blue line).
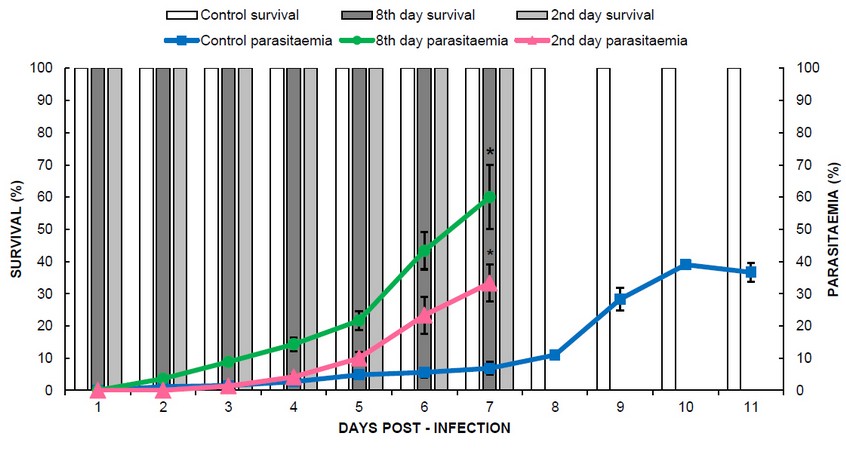
Figure 2. Percentage of parasitaemia over time and survival of control and pregnant mice inoculated on the 2nd and 8th day of gestation. Asterisks (*) indicate significant differences according to ANOVA and Dunnet a posteriori test (p <0.001).
A difference was also evident concerning the days of survival, which are represented with a white bar for the control group, light gray for the group infected at the beginning of pregnancy (2nd day), and dark gray for the group of mice infected at the middle of pregnancy (8th day of gestation). Mice in the control group survived until day 11 post-infection, while mice inoculated on the second and eighth day of gestation died on day 7 post-infection. However, in the control group, the highest parasitaemia was 39% on day 10 post-infection.
Figure 3A shows fetal length in pregnant mice with 9 to 12 fetuses per placenta. Regarding the length of the fetuses, it was affected by infection with P. berghei, and a decrease in the median was observed, corresponding to the fetuses where the mothers were inoculated on the second day of pregnancy (H = 16.04; p ≤ 0.001). The median fetal length in control fetuses was 1.5 cm, while fetuses from mice inoculated on the 2nd-day of pregnancy had a median fetal length of 1.2 cm, with a range of 0.9 to 1.5 cm.
Similarly, the fetus's weight was affected by infection with P. berghei (Figure 3B). The control group weighed 3.1 g, higher than the median of 2.6 g (H = 16.76; p ≤ 0.001). However, the results showed no significant differences in fetal weight between inoculation day 8 and controls. Therefore, there is a marked influence on the effect of infection during pregnancy and the fetus's size.
Figure 3C shows images of fetuses from the three study groups. Apparent differences in the development of mouse fetuses can be observed between infected and non-infected pregnant mothers. The fetuses of mothers infected at the beginning of pregnancy have size and weight obviously below that of fetuses from the control group. The poor development of the hind limbs was also evidenced in comparison to the other two groups of fetuses of the mothers studied.
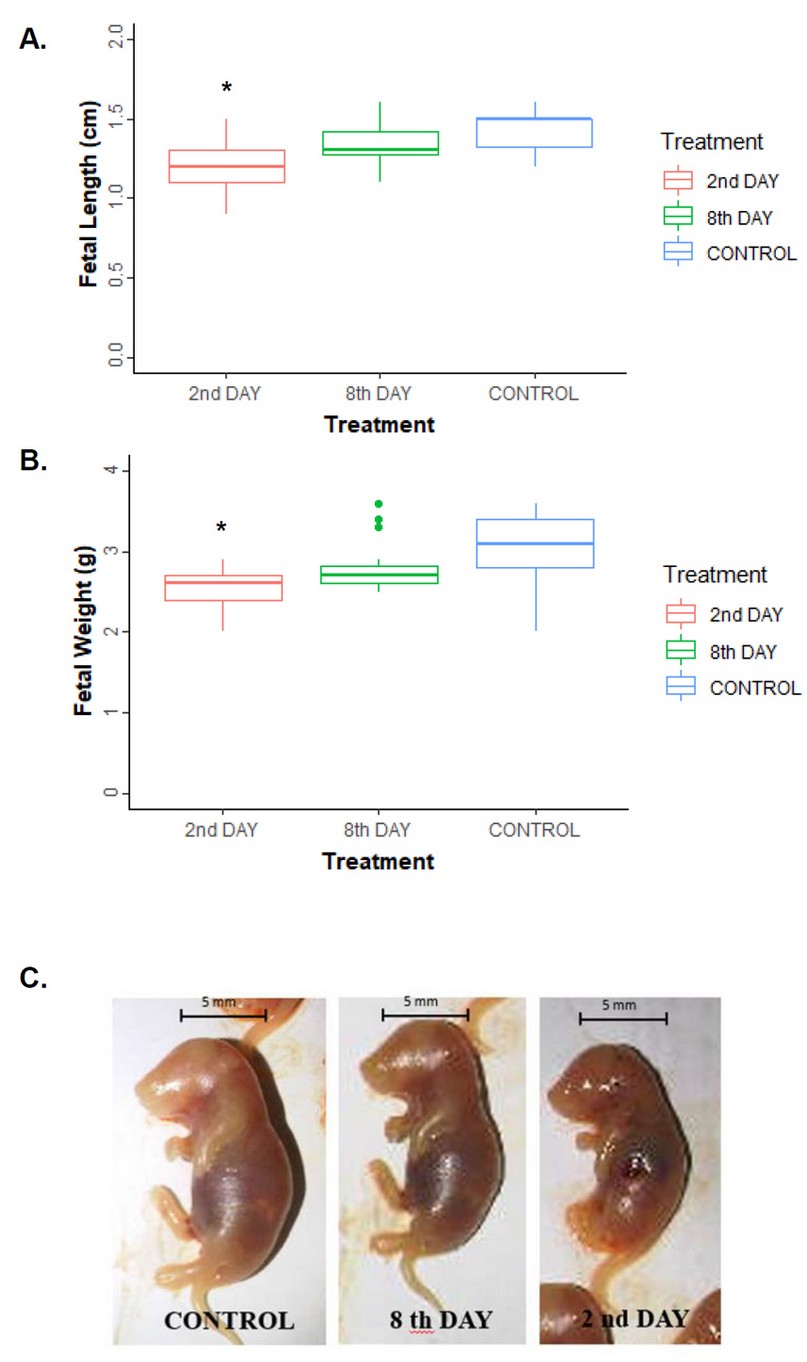
Figure 3: Length (A) and weight (B) of fetuses in control and pregnant mice inoculated with P. berghei on the 2nd and 8th day of gestation. Asterisks (*) indicate significant differences according to Kruskall-Wallis (p <0.001). (C) Representative images of mouse fetuses at 9 days of gestation, which correspond to 7 days post-infection. The line corresponds to a 5 mm scale bar.
Consequently, the time at which malarial infection occurs during pregnancy is critical. The earlier in the pregnancy the infection occurs, the more significant the fetus's negative effects as evidenced by the results of weight, size, and development of the lower extremities.
DISCUSSION
The murine model has always been a valuable tool to evaluate the pathogenicity of malaria infection 1,10. Especially in the mother-fetus binomial, which for ethical reasons is very difficult to evaluate in humans. In the current work, we studied the time during pregnancy when infection by Plasmodium berghei is riskier for the fetus. This evidence will be helpful to in planning and implementing potential passive antibody therapy.
At the beginning of the infection, the parasitaemia curve was similar between mice infected on the second day of pregnancy and control non-pregnant mice at 4 days post-infection. On the contrary, pregnant mice infected on the eighth day of the gestational period developed a more significant infection from day 2 post-infection and reached a maximum parasitaemia of 60%. In comparison, the infected group on day 2 of pregnancy reached the maximum parasitaemia of 33% at 7 days post-infection. The control group survived until day 11 post-infection, and the maximum percentage of parasitaemia was 39% on day 10 post-infection (Figure 2). Pregnant mice are more susceptible to malarial infection, probably because their immune system is suppressed19.
Based on the reports of clinical studies with pregnant mothers, such as Dobbs and Dent24, who analyze the effect of antibodies on fetuses in pregnant women at the end of pregnancy and in the first years of the life of babies. They report that maternal antibodies effectively protect fetuses in the last trimester of pregnancy from infections against the parasite and babies in the first 6 months of life. Other studies in the 1960s showed that the effective recovery of children hospitalized by passive transfer of adult sera caused malaria25,26.
Antibodies can block sporozoites to prevent the invasion of hepatocytes and merozoites to erythrocytes. Thus, as the opzonization of the infected erythrocytes and the Complement system's activation by the VSA molecules expressed on the infected erythrocytes (schizont stage). These antibodies recognize VSA and block interaction with its receptor in endothelial tissue of the placenta mediate the optimization process by monocytes and macrophages, which can engulf parasitized erythrocytes27.
Malaria pregnancy has fatal consequences for both the fetus and the mother. The fact that parasite antigens can cross the syncytiotrophoblast barrier in the last trimester has resulted in the process of immunological tolerance, and consequently an inadequate immune response in the first years of life28,29. Therefore, poor immune response in the first months of life is reported to malaria in pregnancy.
Other studies by Megnekou and co-workers have shown in the BALB/c mice model a correlation between repeated infections of the parasite during pregnancy and concluded that cytokines such as IL-5, IL-12/70, IL -13, and IFN-γ are high. However, low levels of these cytokines were related to anemia and complicated pregnancies. At the same time, that high concentration of IL-10 levels in these pregnancies is correlated with anemia and complicated pregnancies due to the immune-suppression condition20.
Consequently, the moment of infection is crucial in the fetus's development, as can be seen in figure 3, affecting the size and weight of the fetus 21. Our results demonstrate that if the infection occurs early in pregnancy, there is a significant effect on the size reduction and LBW (Figures 3A and B). Reduced limb development was observed in the hind limbs of fetuses from pregnant and infected mice in early gestation (Figure 3C). Unlike this study, Akhabue and colleagues carried out the infection of pregnant mice at the end of the pregnancy on day 13 of gestation. They evaluated the litter's motor development and concluded that they presented changes in their behavior as anxiety and obesity. Nevertheless, it did not affect pups' locomotion when infection occurred during the final gestational period 22. On the other hand, our results support the studies by Huynh and colleagues showing that early intake of sulfadoxine-pyrimethamine in pregnant women reduces the birth of children with low weight; this is correlated with our results infection at the beginning of pregnancy23.
Hence, early treatment of the disease at the beginning of pregnancy by antimalarial drugs is recommended to avoid adverse effects in pregnancy and newborns.
However, fear of teratogenic effects restricts antimalarial drug options in the first trimester. In consequence of this situation, it is necessary to develop other strategies, such as vaccination of mothers and vector control. In this way, the risk of placental infection in early pregnancy could be reduced30,31.
CONCLUSION
Our results showed that the level of parasitaemia in mice infected with P. berghei during the second day of pregnancy could strongly reduce litter size and pup weight than when infection occurs during the middle of the gestational period.
By knowing the time of pregnancy when malarial infections pose significant risks to the fetus, we could suggest alternative therapies for the mother, such as passive transfer of antibodies at the beginning of pregnancy or the use of antimalarial drugs in pregnant women at the beginning of their pregnancy, as reported by other authors. However, resistance to antimalarial drugs generates a problem in treating the disease, so the search for new drugs is necessary, and their studies in the animal model to evaluate teratogenic effects in the biological system of pregnancy.
Data Availability
The data used to support the findings of this study are included in the article.
Conflicts of Interest
The authors declare no conflict of interest regarding the publication of this paper.
Author's Contributions
Conceptualization, formal analysis of the results and writing the manuscript (LMS), experimental assays and analysis of the results (AG), performer of the statistical analysis (BP), writing the manuscript (LZ).
Funding Statement
This investigation was financed in part by a grant from FONACIT (Fondo Nacional de Ciencia, Innovación y Tecnología, No. 29407-2269), Venezuela and Yachay Tech University, Ecuador.
Acknowledgments
We want to thank the staff of the Simón Bolívar University animal house for their technical assistance. We thank Dr. Juan Carlos Piña Crespo for his critical review of this paper.
REFERENCES
1. Boareto A, Gomes C, Centeno J, Golart J, Vergara F, Salum N, et al. Maternal and fetal outcome of pregnancy in Swiss mice infected with Plasmodium berghei ANKAGFP. Reprod Toxicol. octubre de 2019;89:107-14.
2. World Health Organization. World malaria report 2020: 20 years of global progress and challenges. [cited 2021 8 January]. Available from: https://www.who.int/publications/i/item/9789240015791
3. Ofori M, Ansah E, Agyepong I, Ofori-Adjei D, Hviid L, Akanmori B. Pregnancy-associated malaria in a rural community of ghana. Ghana Med J. marzo de 2009;43(1):13-8.
4. Menendez C, Ordi J, Ismail MR, Ventura PJ, Aponte JJ, Kahigwa E, et al. The impact of placental malaria on gestational age and birth weight. J Infect Dis. mayo de 2000;181(5):1740-5.
5. Mens PF, Bojtor EC, Schallig HDFH. Molecular interactions in the placenta during malaria infection. Eur J Obstet Gynecol Reprod Biol. 1 de octubre de 2010;152(2):126-32.
6. Fried M, Duffy PE. Malaria during Pregnancy. Cold Spring Harb Perspect Med. 1 de junio de 2017;7(6) a025551.
7. Kidima WB. Syncytiotrophoblast Functions and Fetal Growth Restriction during Placental Malaria: Updates and Implication for Future Interventions. BioMed Res Int. 2015;2015:451735.
8. Tuikue Ndam NG, Salanti A, Bertin G, Dahlbäck M, Fievet N, Turner L, et al. High level of var2csa transcription by Plasmodium falciparum isolated from the placenta. J Infect Dis. 15 de julio de 2005;192(2):331-5.
9. McLean ARD, Ataide R, Simpson JA, Beeson JG, Fowkes FJI. Malaria and immunity during pregnancy and postpartum: a tale of two species. Parasitology. julio de 2015;142(8):999-1015.
10. Hviid L, Marinho CRF, Staalsoe T, Penha-Gonçalves C. Of mice and women: rodent models of placental malaria. Trends Parasitol. agosto de 2010;26(8):412-9.
11. Umbers AJ, Aitken EH, Rogerson SJ. Malaria in pregnancy: small babies, big problem. Trends Parasitol. 1 de abril de 2011;27(4):168-75.
12. Rogerson SJ, Hviid L, Duffy PE, Leke RF, Taylor DW. Malaria in pregnancy: pathogenesis and immunity. Lancet Infect Dis. 1 de febrero de 2007;7(2):105-17.
13. Neres R, Marinho CRF, Gonçalves LA, Catarino MB, Penha-Gonçalves C. Pregnancy outcome and placenta pathology in Plasmodium berghei ANKA infected mice reproduce the pathogenesis of severe malaria in pregnant women. PloS One. 13 de febrero de 2008;3(2):e1608.
14. Marinho CRF, Neres R, Epiphanio S, Gonçalves LA, Catarino MB, Penha-Gonçalves C. Recrudescent Plasmodium berghei from pregnant mice displays enhanced binding to the placenta and induces protection in multigravida. PloS One. 20 de mayo de 2009;4(5):e5630.
15. Desai M, ter Kuile FO, Nosten F, McGready R, Asamoa K, Brabin B, et al. Epidemiology and burden of malaria in pregnancy. Lancet Infect Dis. febrero de 2007;7(2):93-104.
16. Barateiro A, Pereira MLM, Epiphanio S, Marinho CRF. Contribution of Murine Models to the Study of Malaria During Pregnancy. Front Microbiol. 2019;10:1369.
17. Mangels R, Young B, Keeble S, Ardekani R, Meslin C, Ferreira Z. Genetic and phenotypic influences on copulatory plug survival in mice. Hered Edinb. 2015;115(6):496-502.
18. Spencer L, Quintana D, Hidalgo L. Evaluation of the inhibition of invasion in vitro of P. yoelii strain with different monoclonal antibodies raised against MSP-119. Rev Bioméd. 2008;19(1):45-51.
19. Megnekou R, Hviid L, Staalsoe T. Variant-specific immunity to Plasmodium berghei in pregnant mice. Infect Immun. mayo de 2009;77(5):1827-34.
20. Megnekou R, Staalsoe T, Hviid L. Cytokine response to pregnancy-associated recrudescence of Plasmodium berghei infection in mice with pre-existing immunity to malaria. Malar J. 1 de noviembre de 2013;12:387.
21. Sharma A, Conteh S, Langhorne J, Duffy PE. Heterologous Infection of Pregnant Mice Induces Low Birth Weight and Modifies Offspring Susceptibility to Malaria. PloS One. 2016;11(7):e0160120.
22. Okojie AK, Rauf K, Iyare E. The Impact of Plasmodium berghei Exposure In-utero on Neurobehavioral Profile in Mice. Basic Clin Neurosci. abril de 2019;10(2):99-107.
23. Huynh B-T, Fievet N, Briand V, Borgella S, Massougbodji A, Deloron P, et al. Consequences of gestational malaria on birth weight: finding the best timeframe for intermittent preventive treatment administration. PloS One. 2012;7(4):e35342.
24. Dobbs, K. R. and Dent, A. E . Plasmodium malaria and antimalarial antibodies in the first year of life. Parasitology. 2016; 143(2):129-38.
25. Cohen, S., Mc, G. I. and Carrington, S. Gamma-globulin and acquired immunity to human malaria. 1961; Nature 192, 733–737.
26. McGregor, I. A. The passive transfer of human malarial immunity. American Journal of Tropical Medicine Hygiene.1964; 13(Suppl.), 237–239.
27. Hill, D. L., Eriksson, E. M., Li Wai Suen, C. S., Chiu, C. Y., Ryg-Cornejo, V., Robinson, L. J., Siba, P. M., Mueller, I., Hansen, D. S. and Schofield, L. Opsonising antibodies to P. falciparum merozoites associated with immunity to clinical malaria. 2016; PLoS ONE 8, e74627.
28. Malhotra, I., Dent, A., Mungai, Wamachi, A., Ouma, J.H, Narum,D.L., Muchiri,E., Tisch, D.J., King,C.L. Can prenatal malaria exposure produce an immune tolerant phenotype? A prospective birth cohort study in Kenya. 2009; PLoS medicine. 2009;6(7): e1000116.
29. Feeney, M.E. The immune response to malaria in utero. Immunol Rev. 2020; 293(1): 216–229.
30. Desai, M., Gutman, J., L'Lanziva, A., Otieno, K., Juma, E., Kariuki S., Ouma,P., Were,V., Laserson, K., Katana, A., Williamson A., Kuile, F. Intermittent screening and treatment or intermittent preventive treatment with dihydroartemisinin-piperaquine versus intermittent preventive treatment with sulfadoxine-pyrimethamine for the control of malaria during pregnancy in western Kenya: an open-label, three-group, randomised controlled superiority trial. Lancet. 2015; 386(10012):2507–2519.
31. Kakuru, A., Jagannathan, P., Muhindo, M.K., Natureeba, P., Awori, P., Nakalembe, M., Opira, B., Olwoch, P., Ategeka, J., Nayebare, P., Clark, T.D., Feeney, M.E., Charlebois, E.D., Rizzuto, G., Muehlenbachs, A., Havlir, D.V., Kamya, M.R., Dorsey, G. Dihydroartemisinin-Piperaquine for the Prevention of Malaria in Pregnancy. N Engl J Med. 2016; 374(10):928–939.
Received: 15 January 2021
Accepted: 30 March 2021
Andreina Gómez2, Beatriz Pernía3, Lizbeth Zamora1 and Lilian M. Spencer *1,2
1 School of Biological Sciences and Engineering, Yachay Tech University, San Miguel de Urcuquí, Ecuador;
https://orcid.org/0000-0002-2756-598X; [email protected]; [email protected];
https://orcid.org/0000-0002-4184-1338 [email protected]
2 Cell Biology Department, Simón Bolívar University, Valle de Sartenejas, Caracas, https://orcid.org/0000-0002-9008-9744 Venezuela; [email protected]
3 University of Guayaquil, Faculty of Natural Sciences, Av. Raúl Gómez Lince s/n y Av. Juan Tanca Marengo, Guayaquil, Ecuador. https://orcid.org/0000-0002-2476-7279; [email protected]
Correspondence should be addressed to Lilian M. Spencer; [email protected]