2022.07.02.10
Files > Volume 7 > Vol 7 No 2 2022
Effect of Enterobacter cloacae toxin on immune cells isolated from Leukemia patients

Inas S. Mohammed 1, Rajwa H. Essa 2, Sussain S. Hussain2, * and Khetam H. Rasool 2,*
1 Department of Biomedical Engineering, Biomechanical Branch, University of Technology-Iraq
2 Department of Biology, College of Science, Mustansiriyah University, Baghdad-Iraq.
* Correspondence: Inas S.Mohammed. e-mail: [email protected]; 07702988604
Available from: http://dx.doi.org/10.21931/RB/2022.07.02.10
ABSTRACT
Toxin extract affects Neutrophils and macrophages, which are phagocytic cells. A total of (180) samples, (50) urine and (130) peripheral blood have been obtained from leukemia patients referred to Medical city/ Baghdad Teaching Hospital/ Hematology center; all patients enrolled in this study were diagnosed by the physician according to clinical presentation and laboratory findings as leukemic patients. Investigate the role of Enterobacter cloacae toxins on phagocytic activity/THP-1cells and epithelial line/HBL-100 cells.
THP-1 and HBL-100 cells have been kept in RPMI-1640 that have been supplemented by (10%) of the fetal bovine serum (100µg/mL) of the streptomycin and 100units/mL of the penicillin. The cells have been passaged with the use of the Trypsin-EDTA that has been re-seeded at a confluence of 80% twice every week, and then it has been incubated afterward at a temperature of (37C°). The results of toxin extract from Enterobacter cloacae revealed that (22) fraction/toxins were taken from (11) bacterial samples; these toxins were separated by gel electrophoresis for protein. (18) samples were positive results for gel electrophoresis of protein according to molecular weight: (13.3 KD for α-hemolysin), (70.5 KD for thiol), (75 KD for enterotoxin). This study demonstrates the cytotoxic activity of E. cloacae toxin by using tissue culture through their effect on suppressing the growth of "THP-1 cells" and HBL-100 cells".
Keywords. Tissue culture; Toxin; THP-1 cells; HBL-100 cells; Enterobacter cloacae; Leukemia patients.
INTRODUCTION
Enterobacteriaceae can be defined as a large, heterogeneous set of the Gram (–ve) rods whose natural habitat represents the gastro-intestinal tract of the animals and humans and enteric bacteria, which are referred to as coliforms as well, have a complicated antigenic structure, and create various toxins as well as other factors of virulence. E. cloacae and E. aerogenes took on clinical importance as opportunistic bacteria and appeared as nosocomial pathogens in intensive care patients pathogenic, particularly those on the mechanical ventilations1.
Even though E. cloaca is the major prevalent Enterobacter species responsible for nosocomial infections; not much is identified regarding the factors that influence its virulence and pathogenicity. The pathogenesis may be multifaceted and complex, with the involvement of various putative virulence factors whose significance in disease progression remains unknown1. There are three types of toxin that is produced from Enterobacter species. So, following the adhesion to epithelial cells, in vitro strains of the Enterobacter spp. Create several possible virulence factors, including hemolysin, enterotoxins, and pore-forming thiol-activated cytotoxins similar to the Shiga-like toxin II2. This toxin represents hexamer protein with a (70.5kDa) molecular weight and includes A & B units3. The Thiol-activated cytotoxic toxin created by those bacteria could suppress innate host immune defenses4. Enterotoxin can be defined as a protein exotoxin released by a micro-organism targeting the intestines with a (75kDa) molecular weight. Cytotoxic enterotoxin has been detected through the rounding and the shrinkage of the epithelial cells. The enterotoxins have been often cytotoxic and kill the cells by altering apical membrane permeability of mucosal (i.e., epithelial) cells of the intestinal wall5. Hemolysins are lipids and proteins that cause red blood cell (RBCs) lysis through the cell membrane's disruption. Many hemolysins produced by pathogens cause no considerable destruction of the RBCs during infection6. Cytotoxic activities of the E. cloacae strains have been related to the toxin of (13.3kDa) molecular weight7. The toxin showed stability to the acid treatments at a (2 pH); however, not higher than (7). The hemolysin resulted in the accumulation of the hemorrhagic fluids and the severe histological changes8. Neutrophils and macrophages are phagocytic cells contributing to the primary line related to innate defense against bacterial pathogens through removing and destroying them at the epithelial barrier. To escape the host's innate immune protection, several bacterial pathogens have developed unique ways to decrease the efficient antimicrobial immune response of the macrophages9.
The present study aimed to investigate the role of E. cloacae and their toxins (crude and partially purified toxin) on phagocytic activity (epithelial cell/HBL-100 and macrophage "monocytic cell line"/THP-1).
MATERIALS AND METHODS
Patients
The sample was taken from leukemia patients between (the 14 - > 54) age group in Medical city/ Baghdad Teaching Hospital/ Hematology center, Iraq, from April 2020 to April 2012. The identification of bacterial isolates was to the level of species using the traditional morphological and biochemical tests, according to the methods of McFadden. Confirmed identification with 16S rRNA & type three secretion system. This study showed that E. cloacae had a higher percentage in ALL patients (42.7%) than AML patients (36.6%), and a lesser percent was in CLL patients (11.6%) and CML patients as (8.8%). Furthermore, E. cloacae are one of the opportunistic pathogens11. Comparative efficacy of tissue culture for bacteria and their toxin for detecting Enterobacter cloacae infection.
Bacterial Isolation from blood of Leukemia patients
Leukemia patient's blood of (2-3) ml was taken in test tubes that contained sterile Brain-Heart Infusion medium for blood culturing to ensure the existence of bacteria in the blood samples; it was then left in the incubator at (37C°) for (24h.) for enhancing bacterial growth12. Therefore, all media have been prepared based on the manufacturer's instructions; Trypticase soy agar or MacConky agar were sterilized through autoclaving for (15 minutes), after cooling the media to (56 C°), after incubation on Brain-Heart Infusion for a period of 24 hr in 37C° the suspected positive samples were subculturing on MacConky agar. Otherwise, the medium was sterilized by incubation for (24) hours at (37C°). Pure strains suitable for characterization, the bacterial strains found in positive blood cultures were purified on a Nutrient agar medium. Furthermore, single bacterial strains were recognized using various techniques, including biochemical testing and molecular methods13.
Preparation of cell-free culture filtrate (toxin extraction)
To produce crude toxin, the strains have been propagated in (50ml) of the tryptone soy broth that has been supplemented by (0.6%) extract of the yeast. They were incubated at a temperature of (37C°) with agitation at (120 rpm) in an incubator shaker for (24hour). Cultures have been centrifuged after that in the sterile centrifugation tubes at (10,000rpm) for (30min) at a temperature of (4C°). The resulting supernatant fluids have been filtered via sterile (0.45μm) membrane filters. The cell-free filtrates have been stored at a temperature of (4C°) for not less than (48hour) to be used in the toxin bio-assay14.
Determination of protein concentration
The concentration of the protein has been characterized based on Bradford, with the use of the bovine serum albumin standard curve by the following steps: various concentration values (0, 0.2, 0.4, 0.6, 0.8 and 1 mg/mL) have been made from the (BSA) stock solution (1mg/1ml), after that, (2.50ml) of the prepared Coomassie brilliant blue G250 dye has been added, and then mixed and left to sit for (2min) at the temperature of the room. The absorbance has been measured at (595nm); a blank has been made from (0.45ml) of (0.05 M) phosphate buffer pH7 and (2.5ml) of dye reagent. The standard curve has been plotted between concentrations of the BSA towards corresponding bovine serum albumin absorbance at (595nm).
The concentration of the protein has been calculated through the mixing of (0.05 ml) of the testing sample, (0.45 mL) of the phosphate buffers and (2.5mL) of the Coomassie brilliant blue G250, which has been left to sit for (2min) at the temperature of the room before the measurement of absorbance at (595nm).
Purifications through the chromatography of gel filtration (1x30cm)
Sephadex G1500 has been produced as Pharmacia Fine Chemicals Co has recommended it. A Sephadex G150 quantity has been suspended in (0.05M) of the phosphate buffer pH 7, which has been subjected to the heating at a temperature of (90 C°) for (5h.) to ensure swelling of beads, packed and degassed in a glass column (1x30cm), equilibrated after that with the same buffer. The concentrated sample obtained from the preceding step has been applied to the column. The elution has been accomplished at a (30 ml/h) flow rate, and this same buffer has been utilized for the equilibration. The absorbance of every one of the fractions was assessed at (280 nm). The peak fraction showed toxin activity was collected and concentrated by dialysis against polyethylene glycol for further tests14.
Protein activity has been determined in every one of the fractions, and protein concentration has been determined using Bradford.
Sample preparation and identification
A sample of the protein has been made from the mixing of the protein (6volume) with (1 volume) 0.002% of the BPB (i.e., Bromophenol Blue). All the steps that have been involved in preparing the native protein samples have been carried out at (4 C°). Protein samples for the SDS-polyacrylamide have been substituted in (0.0625M) Tris-HCl (pH 6.8) is brought after that to 5% (v/v) 2-mercaptoethanol, 2% (w/v) SDS, 0.002% (w/v) BPB and 10% (v/v) glycerol utilizing the concentrated stock solution. The samples for the SDS-PAGE have been heated in a bath of boiling water for (3 min), which ensures protein denaturation post-heating. The sample has been left to cool down to the room's temperature (25C°) and centrifuged at (10000 rpm for 5min) 16.
Sample loading and electrophoresis
The approach of the liquid sample loading has been summarized for the native PAGE as well as the SDS-PAGE. Gels have been fixed on the electrophoresis set; the reservoir buffer has been added to the upper line. The sample has been loaded carefully on the surface of the gel with the use of micropipette or a micro syringe. The electrophoresis device was connected to a power pack, and the gel was subjected to (100V) for (2hour) 16.
Gel staining
The Coomassie brilliant blue G250 solution has been made through the dissolution of (100 mg) coomassie blue in (50 ml) a mixture of the Methanol: Water: Glacial acetic acid (5:5:2vol.) with the stirring and its volume has been filled to (1 L) with the distilled water and filtered via Whatman No1 filter paper and kept in the dark bottle at a temperature of (4C°) to be used later.
Tissue culture for Enterobacter cloacae toxin as Table 1.
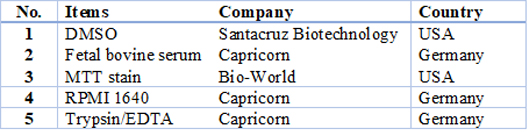
Table 1. Chemicals and reagents
Maintenance of cell cultures
THP-1 and HBL-100 cells were kept in RPMI-1640 supplemented by (10%) of the fetal bovine serum (100µg/mL) of the streptomycin 100units/mL of the penicillin. The cells have been passaged using the Trypsin-EDTA that has been re-seeded at a confluence of 80% twice every week; then, it has been incubated afterward at a temperature of (37C°)17,18.
Cytotoxicity assays
To determine the cytotoxic effects of (Toxin), the assay of the MTT has been done using the (96-well) plates17, 18. The cell lines have been seeded at (1x104cells/well), (24 h.) later, or after achieving a confluent monolayer, the cells have been treated by the tested compounds at a variety of the concentrations. The viability of the cells was measured (72 h.) after the treatment with the removal of the medium, addition of (28.0µL of 2mg/mL) solution of the (MTT) and cell incubation for (2.5 h.) at a temperature of (37C°).
Following the removal of (MTT) solution, crystals that remain in wells have been solubilized by adding (130µL) of the (DMSO) then (37oC) incubation for (15min) with the shaking 19. Absorbency has been specified on the micro-plate reader at 492nm; the assay has been carried out in triplicates. The cell growth inhibition rate (i.e., the cytotoxicity percentage) has been computed based on the following formula20, 21:
(A) represents the control's optical density, and (B) illustrates the samples' optical density21,22.
To visualize the shape of the cell under an inverted microscope, cells were seeded in (24-well) micro-titration plates at (1x105) cells/mL density and incubated for (24 h.) at a temperature of (37C°). After that, the cells were exposed to (Toxin) at a concentration (IC50) for (24 hours). After the exposure, plates were stained with the crystal violet stain and then incubated at (37C°) for (10 –15 min) 22, 23. This stain has been gently washed off using the tap water to the point where the dye has been removed entirely. Cells have been examined using the inverted microscope at a magnification of (40x), and images have been taken using a digital camera attached to a microscope18.
Statistical analysis
The data that were obtained have been analyzed statically with the use of unpaired t-tests by Graph-Pad Prism 617. The values have been given in the mean ± SD of the triplicated measurements24.
RESULTS
Sample preparation and identification
The 30 (23.07%) blood samples were positively cultured using MacConky agar medium as shown in fig.1; isolates were further identified by biochemical tests (table 2). All the isolates were found in smooth form by biochemical tests. Oxidase, H2S, urease and indol were negative, citrate and catalase were positive, and all were found motile. All (30) isolates were found positive by biochemical tests, as shown in Figure 1 and Table 1.
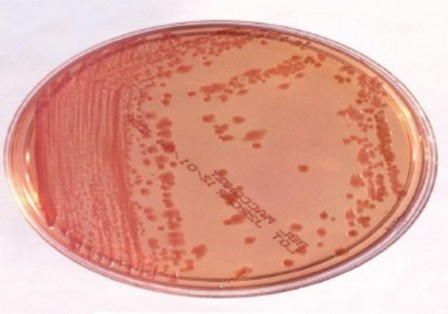
Figure 1. Enterobacter cloacae culture on MacConky agar
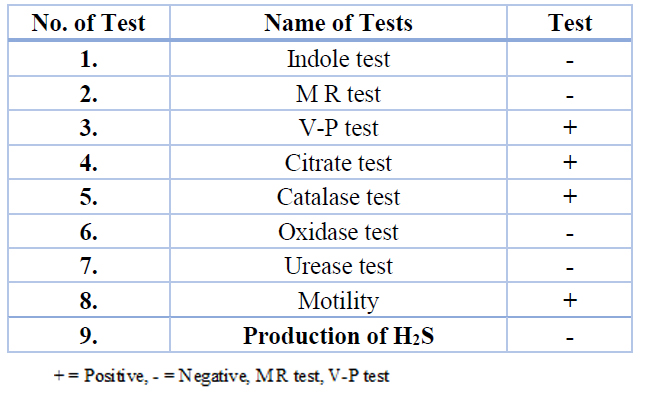
Table 2. Biochemical characters of Enterobacter cloacae isolates
Toxin extraction and protein electrophoresis for Enterobacter cloacae
The isolates were reconfirmed by molecular technique, including 16SS rRNA and TTSS from (11) bacterial samples of E. cloacae we screened for toxin after extraction it, so the result showed that 22 fractions (crude toxin, determination of protein concentration and partially purified toxin) as Figure 2, Figure 3, Fig.4 and Table 3.
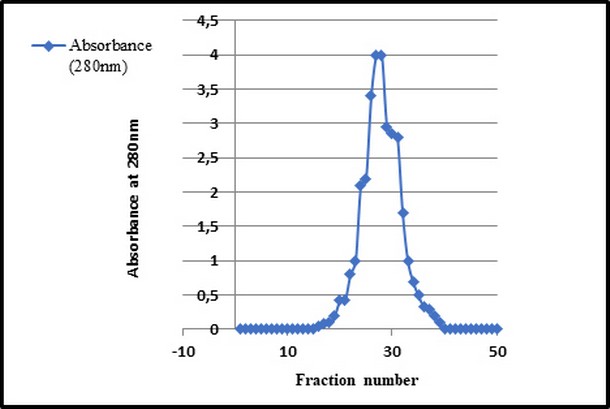
Figure 2. Gel filtration chromatography used Sephadex G 1500 column equilibrated with (0.05 M) sodium phosphate buffer (pH 7.0) at flow rate of (30 ml / hour).
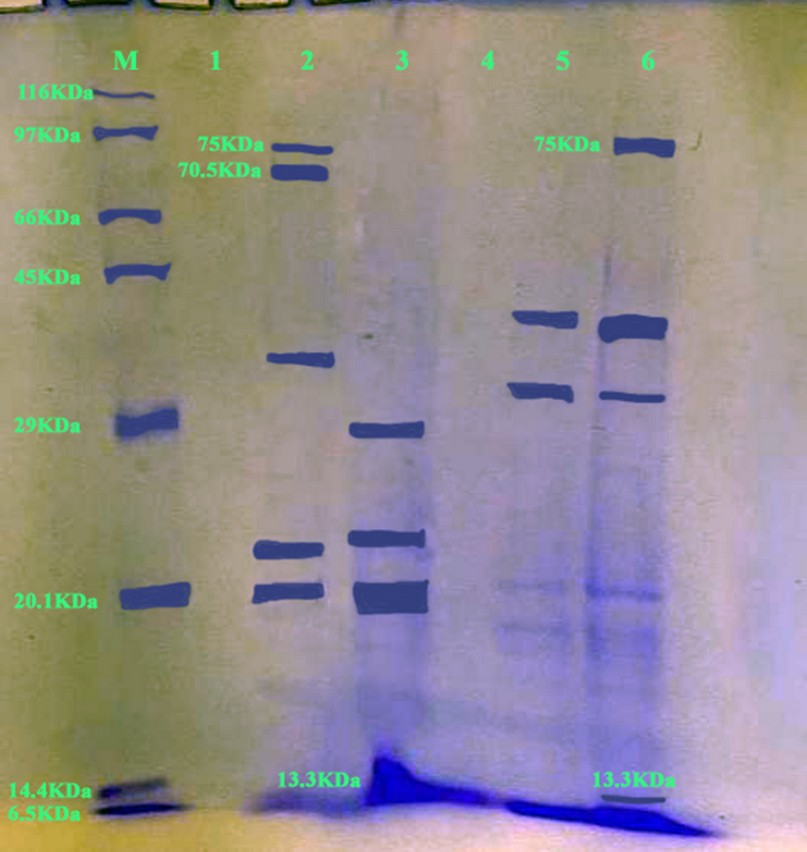
Figure 3: SDS-PAGE gel electrophoresis (1.4gram SDS-PAGE, 100 V/Amp) and Bromophenol Blue dye staining to detect 244 protein "toxin" size product (band 13.3KDa, 70.5 KDa & 75 KDa). Line 2&6 were positive samples and other have negative. M 245 lane: molecular weight confirmation of protein (Marker).
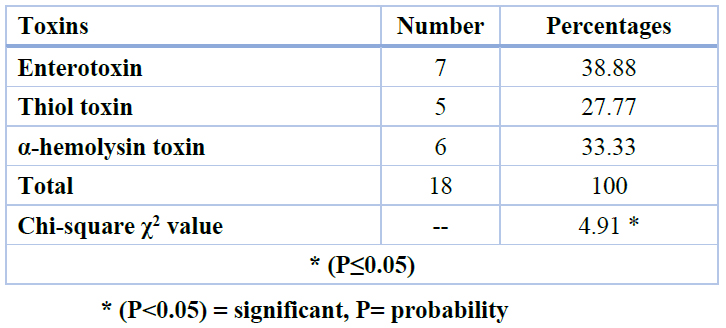
Table 3. Detection of types of toxins for E. cloacae depends on molecular weight using SDS PAGE.
Tissue culture for Enterobacter cloacae toxin
According to toxin extract from Enterobacter cloacae, our results revealed that (22) fraction/toxin were taken from (11) bacterial samples; these toxins were separated by gel electrophoresis for protein. (18) samples were the positive result for gel electrophoresis of protein according to molecular weight: (13.3 KD for α-hemolysin), (70.5 KD for thiol) and (75 KD for enterotoxin). The cytotoxic activity related to E. cloacae strains has been associated with the toxin of α-hemolysin toxin (13.3 kDa) around 6 (33.33%) toxin samples of the isolates26. After isolates of E. cloacae from the blood of leukemia patients and it is culturing on blood agar, it showed that E. cloacae produce α-hemolysin exerted an effect on blood agar through partial lysis regarding red blood cells the disruption of cell membrane for producing a brownish or greenish-grey discoloration around colony as Figure 5.
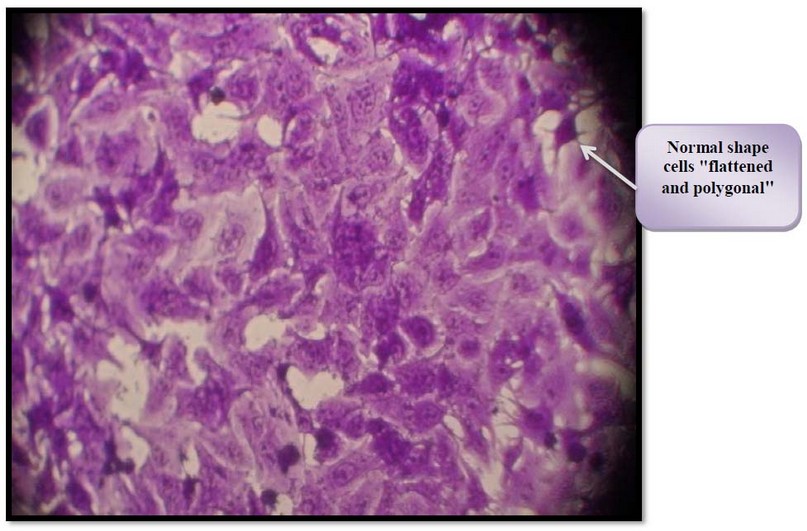
Figure 5. Control untreated HBL-100 cell. Cells
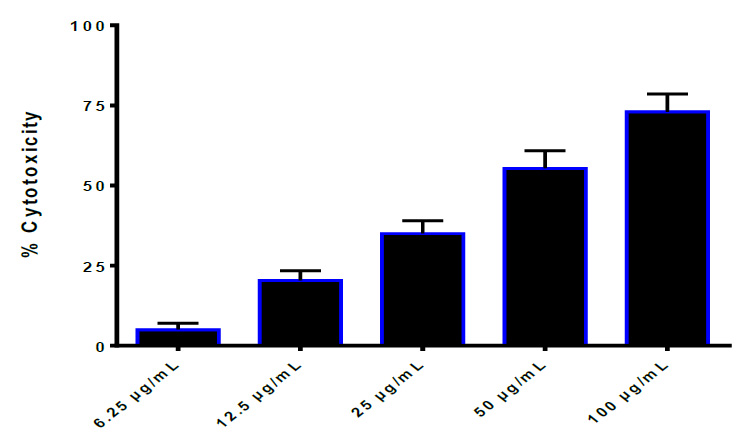
Figure 4. Growth inhibition of HBL-100 cell line effected by thiol. IC50=35.44 μg/ml
The cytotoxic effect of Enterobacter cloacae toxin against THP-1, and HBL-100 cells was studied, as shown in Figures 6 - 13.
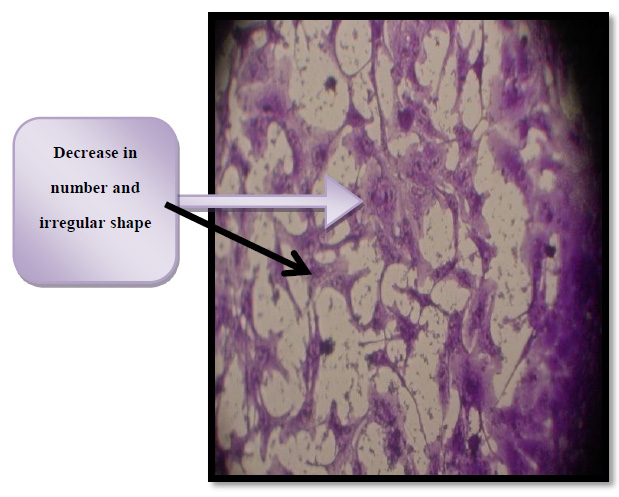
Figure 6. Cytotoxic effect of thiol activated and enterotoxin in HBL - 100 cells. IC50= 37.45 µg/ml
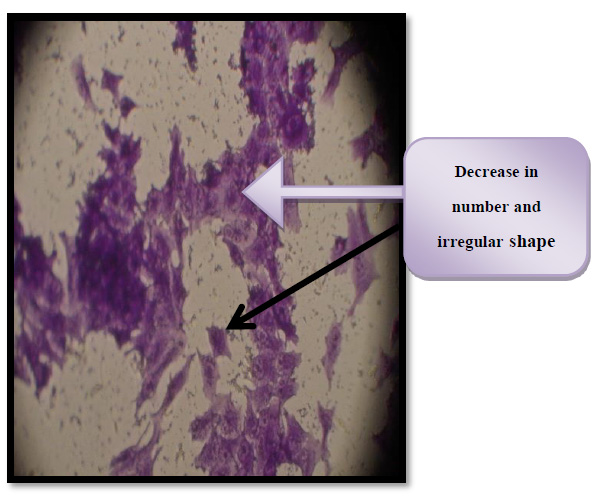
Figure 7 Control untreated HBL-100 cell
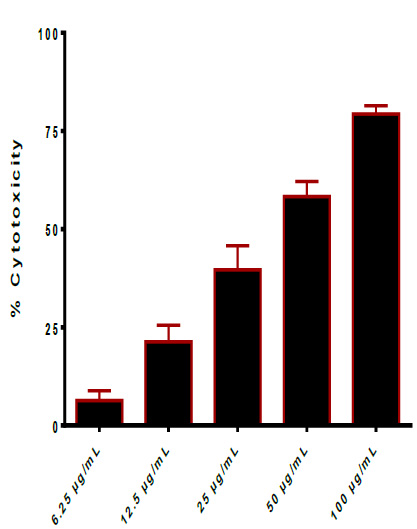
Figure 8. Cytotoxic effect of thiol-activated and enterotoxin in THP-1 cells. IC50= 30.02 μg/ml
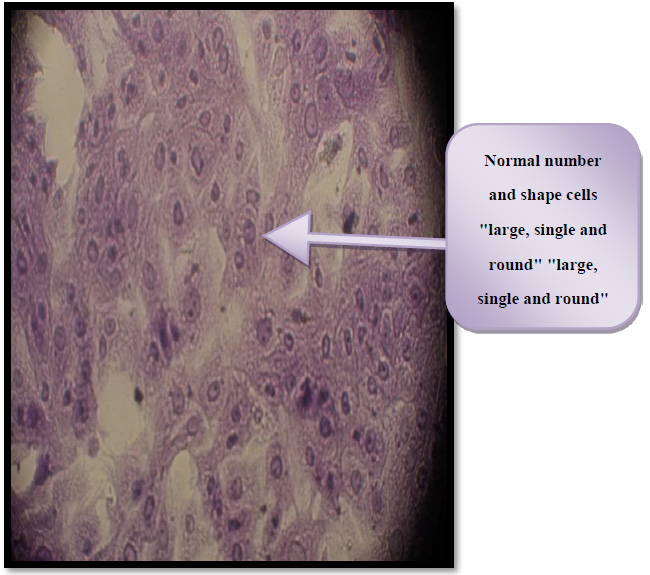
Figure 9. Control untreated THP-1 cells
The cells of both figure (8&9) observed under an inverted microscope at100× magnification.
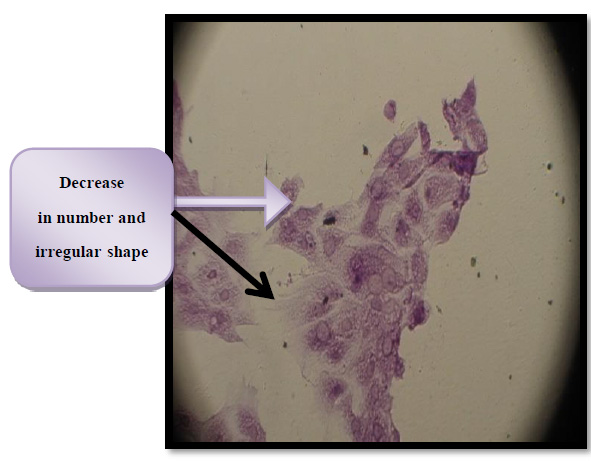
Figure 10. Morphological changes in THP-1 cells affected by thiol toxin activity IC50 for 24 hr
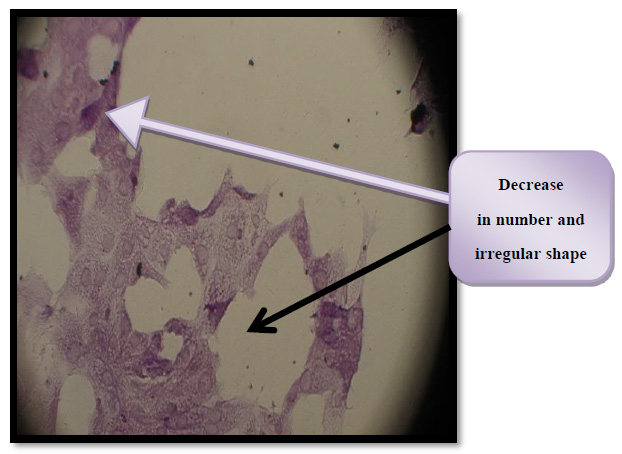
Figure 11. Morphological changes in THP-1 cells affected by enterotoxin activity IC50 for 24 hr.
DISCUSSION
The analyses of incidence that are related to the markers of virulence in the isolates of E. cloacae indicated that 18 (81.81%) produced cytotoxic toxins and the absorbency of toxin has been determined on a microplate reader at (595 nm) for crude was (0.8 nm) and for toxin was (0.2 nm). The activity was observed on macrophage and epithelial cells. Cytotoxic strains were detected from blood leukemia patients. After that used gel electrophoresis of protein, showed 18 (81.81%) samples contain bacterial toxin out of (22) fraction and it was separated by molecular weight.
The cytotoxic activity related to E. cloacae strains has been related to the toxin of α-hemolysin toxin (13.3 kDa)25. After isolates of E. cloacae from blood of leukemia patients and it culturing on the blood agar, showed E. cloacae produce α-hemolysin exerted an effect on blood agar through partial lysis regarding red blood cells through the disruption of cell membrane for producing a brownish or greenish-grey discoloration around colony. Those monomers are diffused to target cells and attached to them through certain receivers. After that, they oligomerize, resulting in ring-shaped heptamer complexes26. Rapid discharges of essential molecules like the ATP, the dissipation of ion gradients and membrane potential, and irreversible osmotic swelling resulting in cell wall rupture (i.e. lysis) might all contribute to hosting cell death27.
In this study, we have 5 (27.77%) toxin samples of that were cytotoxic to THP-1 and HBL-100. The findings imply that the bacteria's thiol-activated (70.5 kDa) cytotoxic toxin might impair the host's innate immune response. E. cloacae strains28, identified a novel thiol-activated toxin that had an impact on leukocytes and erythrocytes. A high dose of toxin produced leukocyte lysis prior to apoptosis was triggered29. Also, in this work, we have 7 (38.88%) enterotoxin samples of the isolates that cytotoxic to THP-1 and HBL-100.
Enterotoxin (75 kDa) is produced and secreted from E. cloacae. Enterotoxins are typically cytotoxic, killing cells via affecting the permeability of epithelial cells in the intestinal wall's apical membrane. Pore-forming toxins (mainly chloride pores) are bacteria-secreted toxins that combine to produce pores in cell membranes30. The cytotoxic effect of toxin extraction against cell lines (THP-1 and HBL-100) was studied. The capacity of thiol activated and enterotoxin to decrease the proliferation of cell lines was used to investigate their antitumor efficacy (HBL-100 and THP-1). The results of this work revealed significantly considerable cytotoxic activity against human cancer cell lines (HBL-100 and THP-1). The findings reveal that thiol activated and enterotoxin can inhibit the growth of cell lines (HBL-100 and THP-1), and that this impact is concentration-dependent.
In this work, we looked at the virulence factor (toxin) production and associated aspects of the human isolates of the E. cloacae. The cytotoxic effect of bacteria and its toxin on macrophages (THP-1) and epithelial cells (HBL-100) was investigated. Another work found that E. cloacae human isolates produced virulence factor (toxin) related features in Vero, CHO (i.e., Chinese hamster ovary), and murine macrophage J-774 cells and the existence of TTSS31.
CONCLUSIONS
This study focused upon the explanation of contributions of cytotoxic activities, culture and also focused on tissue culture for E. cloacae pathogenesis toxin extraction, and discovered during our study that the tissue culture observed bacterial toxin causes inhibition and suppress to monocytic cell "THP-1 cells" and epithelial cells "HBL-100 cells".
Funding: "Self funded this research.
Informed Consent Statement:
"Written informed consent has been obtained from consultants of hematology center referred to Medical city/ Baghdad Teaching Hospital/ Hematology center.
Data Availability Statement: The results might be achieved using an evaluation software or assessment table, which could be available at www.diagnostics.sk/idmicro.
Acknowledgments: This includes personal support "self-financing" (e.g., materials used for experiments).
Conflicts of Interest: None
REFERENCES
1. Mezzatesta, M. L.; Gona, F.; Stefani, S. Enterobacter cloacae complex: clinical impact and emerging antibiotic resistance. Future Microbiol. 2012, 7:887-902. doi: 10.2217/fmb.12.61.
2. Barnes, A. I.; Paraje, M. G.; Del, C.; Battan, P.; Albesa, I. Molecular properties and metabolic effect on blood cells produced by a new toxin of Enterobacter cloacae. 2001, 17: 409–418.
3. Falguieres, T.; Maak, M.; von Weyhern, C.; Sarr, M.; Sastre, X.; Poupon, M. F. Human colorectal tumors and metastases express Gb3 and can be targeted by an intestinal pathogen-based delivery tool. 2008, Volume 7, pp. 2498-2508.
4. Paraje, M. G.; Barnes, A. I.; Albesa, I. An Enterobacter cloacae toxin able to generate oxidative stress and to provoked dosedependent lysis of leucocytes. Int. J. Med. Microbiol. 2005, 295: 109–116.
5. Dinges, M. M.; Orwin, P. M.; Schlievert, P. M. Exotoxins of bacteria. Clinical Microbiology Reviews. 2000, 13 (1): 16–34, table of contents. doi:10.1128/CMR.13.1.16. ISSN 0893-8512. PMC 88931. PMID 10627489.
6. Vesper, S. J.; Vesper, M. J. Possible role of fungal hemolysins in sick building syndrome. Adv. Appl. Microbiol. Advances in Applied Microbiology. 2004, 55. pp. 191–213. doi:10.1016/S0065-2164(04)55007-4. ISBN 9780120026579. PMID 15350795.
7. Albesa, I.; Barbnes, AI.; Paraje, M. G. Induction of oxidative stress in leukocytes by an Enterobacter cloacae toxin able to form oligomers and binding to proteins. Biochem Bioph Res. 2000, Co 274: 649–654.
8. Chalmeau, J.; Monina, N.; Shin, J.; Vieu, C.; Noireaux, V. α-Hemolysin pore formation into a supported phospholipid bilayer using cell-free expression. Biochim. Biophys. Acta. 2011, 1808 (1): 271–8. doi:10.1016/ j.bbamem.2010.07.027. PMID 20692229.
9. Krzyminska, S.; Mokracka, J.; Koczura, R.; Kaznowski, A. Cytotoxic activity of Enterobacter cloacae human isolates. FEMS Immunol. Med. Microbiol. 2010, 56, 248–252.
10. McFadden, J. F. Biochemical tests for identification of medical bacteria, 3rd ed; Lippincott Williams and Wilkins, USA, 2000
11. Kamal, A. S. Risk of Bacteremia and Fungemia in Children with Acute Lymphoblastic Leukemia and Febrile Neutropenia. Benha University. 2010.
12. Nicolas, B.; Michael, D.; Isaline, C.; Jean-Paul, F.; Stephane, L.; Nadir, A.; Benoit, S.; Elie, A. Diagnostic Accuracy of Procalcitonin in Critically Ill Immunocompromised Paients. BMC Infectious Diseases. 2011, 11 (224), 1-6.
13. Galanta, Slovakn Republic Company. 2019.
14. Trower, C. J.; Abo, S.; Majeed, K.; voni Itzsteyn, M. Production of an Enterotoxin by a gastro-entertis associated Aeromonas strain. J Med. Microbiol. 2000, 49,121-126.
15. Bradford, M. A rapid and sensitive method for the quantitationof microgram quantities of protein using the principle of protein-dye binding. Anal. Biochem. ,1979, 72: 248-254.
16. Hames, B.D. An introduction to polyacrylamide gel electrophoresis of proteins Dlm. B. D. Hames and D. Richwood (pnyt.). Ed. Gel electrophoresis of proteins: a practical approach hlm. 1-91. Oxford: IRL Press, 1985.
17. Khashan, K. S.; Sulaiman, G. M.; Hussain, S. A.; Marzoog, T. R.; Jabir, M. S. Synthesis, Characterization and Evaluation of Anti-bacterial, Anti-parasitic and Anti-cancer Activities of Aluminum-Doped Zinc Oxide Nanoparticles. Journal of Inorganic and Organometallic Polymers and Materials. 2020, 1-17.
18. Jabir, M.; Sahib, U. I.; Taqi, Z.; Taha, A.; Sulaiman, G.; Albukhaty, S.; Rizwana, H. Linalool-Loaded Glutathione-Modified Gold Nanoparticles Conjugated with CALNN Peptide as Apoptosis Inducer and NF-κB Translocation Inhibitor in SKOV-3 Cell Line. International Journal of Nanomedicine, 2020, 15, 9025.
19. Al-Ziaydi, A. G., Al-Shammari, A. M., Hamzah, M. I., Kadhim, H. S.; Jabir, M. S. Newcastle disease virus suppress glycolysis pathway and induce breast cancer cells death. VirusDisease. 2020, 1-8.
20. Khashan, K. S.; Jabir, M. S.; Abdulameer, F. A. Carbon Nanoparticles prepared by laser ablation in liquid environment. Surface Review and Letters, 2019, 26(10), 1950078.
21. Kareem, S. H.; Naji, A. M.; Taqi, Z. J.; Jabir, M. S. Polyvinylpyrrolidone Loaded-MnZnFe 2 O 4 Magnetic Nanocomposites Induce Apoptosis in Cancer Cells Through Mitochondrial Damage and P 53 Pathway. Journal of Inorganic and Organometallic Polymers and Materials. 2020, 1-15.
22. Jabir, M. S.; Hussien, A. A.; Sulaiman, G. M.; Yaseen, N. Y.; Dewir, Y. H.; Alwahibi, M. S. ; Rizwana, H. Green synthesis of silver nanoparticles from Eriobotrya japonica extract: a promising approach against cancer cells proliferation, inflammation, allergic disorders and phagocytosis induction. Artificial Cells, Nanomedicine, and Biotechnology. 2021, 49(1), 48-60.
23. Waheeb, H. M.; Sulaiman, G. M.; Jabir, M. S. Effect of hesperidin conjugated with golden nanoparticles on phagocytic activity: In vitro study. In AIP Conference Proceedings. 2020, Volum 2213, No. 1, pp. 020217.
24. Al-Shammari, A. M.; Al-Saadi, H.; Al-Shammari, S. M.; Jabir, M. S. Galangin enhances gold nanoparticles as anti-tumor agents against ovarian cancer cells. In AIP Conference Proceedings. 2020, Volume 2213, No. 1, pp. 020206.
25. Majid, S.; Jabir; Yasmin, M.; Saleh, Ghassan, M.; Sulaiman, Nahi Y.; Yaseen, Usama, I.; Sahib; Yaser Hassan Dewir; Mona, S.; Alwahibi; Dina A. Soliman Green Synthesis of Silver Nanoparticles Using Annona muricata Extract as an Inducer of Apoptosis in Cancer Cells and Inhibitor for NLRP3 Inflammasome via Enhanced Autophagy. Nanomaterials. 2021, 11, 384. 1-22.
26. Sameen, A. M.; Jabir, M. S.; Al-Ani, M. Q. Therapeutic combination of gold nanoparticles and LPS as cytotoxic and apoptosis inducer in breast cancer cells. In AIP Conference Proceedings. 2020, Volum 2213, No. 1, pp 020215.
27. Albesa, I.; Barbnes, AI.; Paraje, M. G. Induction of oxidative stress in leukocytes by an Enterobacter cloacae toxin able to form oligomers and binding to proteins. Biochem Bioph Res. Co. 2000, 274: 649–654.
28. Thompson, J. R.; Cronin, B.; Bayley, H.; Wallace, M. I. Rapid assembly of a multimeric membrane protein pore. Biophys. J. 2011, 101 (11): 2679–83. Bibcode: 2011BpJ...101.2679T. doi:10.1016/j.bpj. 2011.09.054. PMC 3297801. PMID 22261056.
29. Paraje, M. G.; Barnes, A. I.; Albesa, I. An Enterobacter cloacae toxin able to generate oxidative stress and to provoked dosedependent lysis of leucocytes. Int. J. Med. Microbiol. 2005, 295: 109–116.
30. Hornby, P. J. Drug discovery approaches to irritable bowel syndrome. Expert Opinion on Drug Discovery. 2015, 10 (8): 809– 24. doi:10.1517/17460441.2015.1049528. PMID 26193876. S2CI D 207494271.
31. Sylwia, K.; Joanna, M.; Ryszard, K.; Adam K. Immunology & Medical Microbiology. 2009, Volume 56, Issue 3, Pages 248–252.
Received: 7 September2021 / Accepted: 18 December 2021 / Published:15 May 2022
Citation: Inas S. Mohammed.; Rajwa H. Essa; Sussain S. Hussain ; Khetam H. Rasool. Effect of Enterobacter cloacae toxin on immune cells isolated from Leukemia patients. Revis Bionatura 2022;7(2) 10. http://dx.doi.org/10.21931/RB/2022.07.02.10