2022.07.02.3
Files > Volume 7 > Vol 7 No 2 2022
Cambios en la concentración de ácido fítico, fósforo libre y hierro soluble durante la fermentación de repollo blanco y repollo chino
Changes in phytic acid concentration, free phosphorus and soluble iron during fermentation of white cabbage and Chinese cabbage

Romina B. Parada 1,2*, Franco Sosa 1,2, Emilio Marguet 2 and Marisol Vallejo 2
1 Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina.
2 Laboratorio de Biotecnología Microbiana. Facultad de Ciencias Naturales y Ciencias de la Salud (Sede Trelew). Universidad Nacional de la Patagonia. Argentina.
* Correspondence: [email protected]
Available from: http://dx.doi.org/10.21931/RB/2022.07.02.3
RESUMEN
Se estudió la evolución de la concentración de ácido fítico, fósforo libre y hierro soluble durante la fermentación de repollo blanco y repollo chino. En ambos casos, la máxima población de bacterias ácido lácticas se logró a los cinco días del proceso y luego disminuyó continuamente hasta el final. El pH inicial del repollo blanco y repollo chino fue de 6,1 y durante los primeros cinco días disminuyó a 3,7 y 4,3 respectivamente, luego permanecieron estables hasta los 30 días. En el repollo blanco, la concentración de ácido fítico disminuyó y el fósforo libre se incrementó durante los primeros cinco días, después no se detectaron cambios significativos. En el repollo chino, la degradación del ácido fítico se observó durante los primeros 15 días, mientras que el fósforo libre aumentó hasta el final del proceso. Ambos vegetales mostraron una concentración inicial de hierro comparable, luego, se observó un incremento hasta el final del proceso, siendo este fenómeno más notable en el repollo chino. Los resultados obtenidos sugieren que la degradación del ácido fítico producida durante la fermentación por la actividad de fitasas vegetales y bacterianas, no sólo origina la liberación de fósforo libre, sino que mejora la bioaccesibilidad del hierro.
Palabras claves. biodisponibilidad de nutrientes, fermentación espontánea, Brassica
ABSTRACT
The evolution of phytic acid concentration, free phosphorus, and soluble iron was studied through the fermentation of white cabbage and Chinese cabbage. In both cases, the maximum population of lactic acid bacteria was achieved at five days of process and then decreased continuously until the end. The initial pH in white cabbage and Chinese cabbage was 6.1, decreased to 3.7, and 4.3, respectively, at five days and remained stable until fermentation. The decrease of phytic acid concentration and the increase of free phosphorous could be observed in white cabbage during the first five days of fermentation; then, both parameters did not show significant changes. In Chinese cabbage, the degradation of phytic acid could be observed during the first 15 days of fermentation, while free phosphorus increased continuously through 30 days. At the beginning of fermentation, both vegetables displayed comparable iron concentration, then the increase could be observed steadily until the end of the process; however, this phenomenon was more remarkable in Chinese cabbage. The results obtained suggest that phytic acid degradation produced during fermentation by the activity of vegetables and bacterial phytases originates the release of free phosphorus and enhances iron bioaccessibility.
Keywords. Nutrient bioavailability, spontaneous fermentation, Brassica.
INTRODUCCIÓN
Las fermentaciones han sido utilizadas históricamente por el hombre con el propósito de aumentar la vida útil de los alimentos y eliminar o inhibir bacterias patógenas o deteriorantes. En el caso específico de la flora epifítica de los vegetales, especialmente las bacterias ácido lácticas (BAL), utilizan los hidratos de carbono presentes en el tejido de las plantas transformándolos en ácidos orgánicos, logrando de esta manera un marcado descenso del pH y, en consecuencia la inhibición del desarrollo de bacterias indeseadas 1. Además de este fenómeno, se producen distintas reacciones metabólicas que influyen por ejemplo, en la capacidad antibiótica mediante la formación de peróxido de hidrógeno, diacetilo y bacteriocinas 2; aumento de la capacidad antioxidantes como consecuencia de la hidrólisis de polifenoles y flavonoides 3,4; incremento de moléculas bioactivas como la producción de tioglicolatos a partir de glucosinolatos por la acción de mirosinasas 5,6, entre otras.
Un cambio importante que se produce durante la fermentación es la disminución o eliminación de factores antinutricionales dentro de los que tenemos que considerar a los inhibidores de enzimas degradativas del tracto gastrointestinal, especialmente la tripsina, o moléculas que interfieren en la biodisponibilidad de nutrientes 7. En este grupo de moléculas uno de los más importantes lo constituye el ácido fítico y sus derivados. Aproximadamente el 80 % del total del fósforo contenido en las células vegetales está presente como fitato y no solo su biodisponibilidad resulta prácticamente inaccesible 8,9 sino que la liberación al medio ambiente podría provocar efectos negativos, como sucede con la eutrofización de los cuerpos de agua en las zonas donde se practica la ganadería y agricultura intensiva10.
El ácido fítico es definido desde el punto de vista químico como ácido mioinositol (1,2,3,4,5,6) hexafosfórico. Su estructura le permite formar quelatos estables e insolubles con cationes divalentes como Ca++, Mg++, Zn++, Cu++ y Fe++, fenómeno que trae como consecuencia una disminución en la absorción y digestión de estos minerales en el tracto intestinal 11. También tienen la capacidad de formar complejos con las proteínas disminuyendo su solubilidad e interfiriendo su digestibilidad 11.
En consecuencia, la degradación del ácido fítico o sus derivados durante el procesamiento de alimentos de origen vegetal es esencial para aumentar la disponibilidad de fósforo y oligoelementos. En el caso específico del hierro, algunos autores afirman que para mejorar sustancialmente su absorción la degradación del ácido fítico deber ser casi completa 12.
Los tratamientos térmicos, la cocción, el malteado y la fermentación son procesos que logran disminuir la concentración de estas moléculas quelantes. En el caso específico de la fermentación, la acción conjunta de las fitasas vegetales y microbianas logran hidrolizar y liberar fósforo soluble a partir de los fitatos 1,13,14.
Durante los últimos años, ha cobrado un significativo interés el estudio de la fermentación de brasicáceas debido a su alto consumo en distintas regiones del mundo y el alto valor nutritivo que poseen 15. Estos vegetales son ricos en carbohidratos, fibras, vitaminas y minerales. Además, su capacidad antioxidante tiene influencia positiva en la lucha contra patologías humanas tales como enfermedades cardiovasculares, cáncer, diabetes, etc 4. Las brasicáceas son cultivadas con relativa facilidad gracias a su adaptación a diferentes climas y calidad de suelos y, en algunos países constituyen la base de su alimentación 15.
El objetivo de este trabajo fue determinar la evolución del contenido de ácido fítico, fósforo y hierro soluble durante la fermentación espontánea de repollo chino (Brassica rapa L. ssp. glabra, Regel) y el repollo blanco (B. oleracea L. ssp. capitata, (L) Metzg) cultivados en el Valle Inferior del Río Chubut (Patagonia, Argentina).
MATERIALES Y MÉTODOS
Proceso de fermentación
El repollo chino (Brassica rapa L. ssp. glabra, Regel) y el repollo blanco (B. oleracea L. ssp. capitata, (L) Metzg) fueron obtenidos de establecimientos localizados en el Valle Inferior del Río Chubut, Patagonia, Argentina (latitud -43,14; longitud -65,19; 11 metros sobre el nivel del mar).
Con el objeto de preparar el material a fermentar se eliminaron las hojas exteriores, los bulbos limpios fueron cortados en tiras de 2 mm de espesor y se colocaron en frascos. Se adicionó NaCl en una concentración final del 3 % (m/m). Ambos vegetales se fermentaron a 18 °C durante 30 días. Las pruebas se realizaron por duplicado.
Parámetros de fermentación
La toma de muestras para determinar el pH y el recuento de BAL se realizó a los días 0, 1, 2, 3, 4, 5, 10, 15, 20, 25 y 30 del proceso de fermentación. La determinación del pH se realizó con un pH-metro calibrado modelo Orion 410. El recuento se realizó en agar Man Rogosa Sharpe (MRS) (Laboratorios Britania, Buenos Aires, Argentina) a 30 °C durante 24-48 h. Los resultados se expresaron en Unidades Formadoras de Colonia por mililitro (UFC/ml).
Determinación de fosfato libre
Las muestras se centrifugaron a 2000 rpm durante 5 min en una centrífuga Rolco CP 2036 (Buenos Aires, Argentina). El fosfato libre se determinó utilizando 100 µl de sobrenadante y 900 µl de una solución que contenía 0,6 M de H2SO4, 2 % (m/v) de ácido ascórbico y 0,5 % (m/v) de molibdato de amonio. Luego de una incubación a 50 ºC durante 30 min se realizó la lectura de la densidad óptica a 820 nm. Para la curva de calibración se utilizó una solución patrón de K2HPO4. Los resultados fueron expresados en mg de fósforo/100 ml 16.
Determinación de ácido fítico
La concentración de ácido fítico en los sobrenadantes se determinó utilizando el kit comercial Megazime Internacional (Wicklow, Irlanda) siguiendo las instrucciones del fabricante. El ensayo se basa en la hidrólisis del ácido fítico por la actividad de la fitasa en mio-inisitol-fosfato y fósforo libre, y la posterior actividad de la fosfatasa alcalina que libera el resto de los fosfatos de la molécula de mio-inositol 17. El fosfato libre se determinó según se describió anteriormente y los resultados se expresaron en mg ácido fítico/100 ml, utilizando la siguiente ecuación:
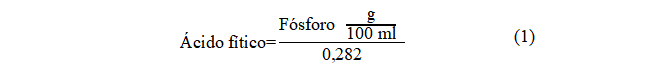
Determinación de hierro libre
Se trataron 200 µl de sobrenadante con 1 ml de una solución 30 mM de ácido ascórbico en buffer citrato (0,05 M, pH 4,5). Luego de una incubación a 37 °C durante 5 min se agregó 200 µl de una solución de ferrozime (5 mM). Se incubó a 37 °C durante 40 min y se midió la densidad óptica a 562 nm en un espectrofotómetro Jenway modelo 6405 (Londres, Reino Unido). Se realizó una curva patrón con Fe(NO3)3. Los resultados se expresaron en mg hierro/100 ml 18.
RESULTADOS Y DISCUSSION
Parámetros de fermentación
El descenso de pH y el recuento de BAL constituyen los parámetros que permiten determinar la evolución del proceso de fermentación. La población inicial de BAL en ambos vegetales fue: 2,1 y 2,5 log UFC/mL para repollo blanco y repollo chino, respectivamente, estos valores coinciden con reportes previos 1. Los vegetales mostraron el máximo de población luego de 5 días de proceso, determinándose un recuento de 9,2 log UFC/mL para repollo blanco y 9,5 log UFC/mL para repollo chino. Posteriormente, la población de BAL comenzó a descender en ambos vegetales, alcanzando un valor próximo a 6,0 log UFC/mL después de 30 días (Figura 1). El repollo blanco exhibió un pH inicial de 6,0 y descendió hasta lograr un valor de 3,7 en el día 10, manteniéndose estable hasta el final del proceso. En el caso del repollo chino, el pH inicial fue de 6,1 y descendió a 4,3 en el día 5, valor que se mantuvo estable hasta los 30 días de ensayo (Figura 1). La marcada diferencia en los valores de pH alcanzados luego de 30 días de experiencia, es atribuible a la mayor concentración de carbohidratos que exhibe el repollo blanco con respecto al repollo chino 19, fenómeno que permite una síntesis mayor de ácidos orgánicos y el consecuente aumento de la acidez titulable.
El descenso de la población de BAL observado, luego de alcanzar el valor máximo, se debe a las limitantes nutricionales de los vegetales estudiados. En el caso particular de las brasicáceas, la baja concentración de proteínas no permite la duplicación celular una vez agotado este recurso. Las BAL, no obstante adaptarse y duplicarse con velocidad en sustratos vegetales, presentan numerosas auxotrofías que las hace dependientes de una constante provisión de proteínas, péptidos y aminoácidos 20.
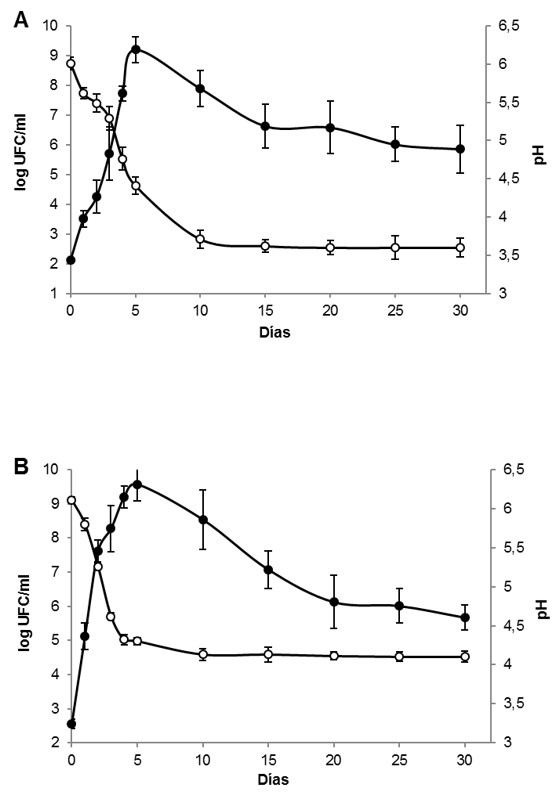
Figura 1. Evolución del pH (○) y población de bacterias ácido lácticas (log UFC/ml) (●) durante la fermentación del repollo blanco (a) y del repollo chino (b).
Determinación de fosfato libre y ácido fítico
El ácido fítico y sus derivados constituyen la mayor reserva de fósforo de las plantas, sin embargo, la mayoría de los animales monogástricos no logran degradarlo por carecer de fitasas o exhibir actividades enzimáticas insuficientes en su mucosa intestinal 8. La degradación de fitatos en los alimentos es de vital importancia para aumentar la biodisponibilidad de nutrientes, ya que evita que estos formen quelatos insolubles en el tracto intestinal 11,21.
Durante la fermentación espontánea, las fitasas vegetales son liberadas al medio degradando el ácido fítico y aumentando la concentración de fósforo soluble. En forma paralela, las BAL aumentan su población y contribuyen a la degradación mediante su actividad fitásica extracelular 22. En los casos estudiados, puede observase que en el repollo blanco la concentración inicial de ácido fítico fue de 145,9 mg/100 ml, descendiendo hasta 101,1 mg/100 ml en el día 5 y manteniéndose estable hasta el final del proceso fermentativo. En forma inversa, la concentración de fósforo soluble aumentó desde una concentración inicial de 1,1 mg/100 ml hasta estabilizarse en el día 5 en 7,4 mg/ 100 ml manteniéndose sin modificaciones hasta los 30 días de experiencia. Mientras que, en el repollo chino se determinó una concentración inicial de ácido fítico de 119 mg/100 ml pero a diferencia del repollo blanco, el descenso se observó hasta el día 15, alcanzado una concentración de 25 mg/100 ml al final de la experiencia. En cuanto a la concentración de fósforo soluble, el repollo chino presentó una mayor concentración inicial (19,4 mg/100 ml) que en el repollo blanco; y el incremento se observó de manera continua durante todo el proceso alcanzando un valor final de 29,6 mg/ml (Figura 2).
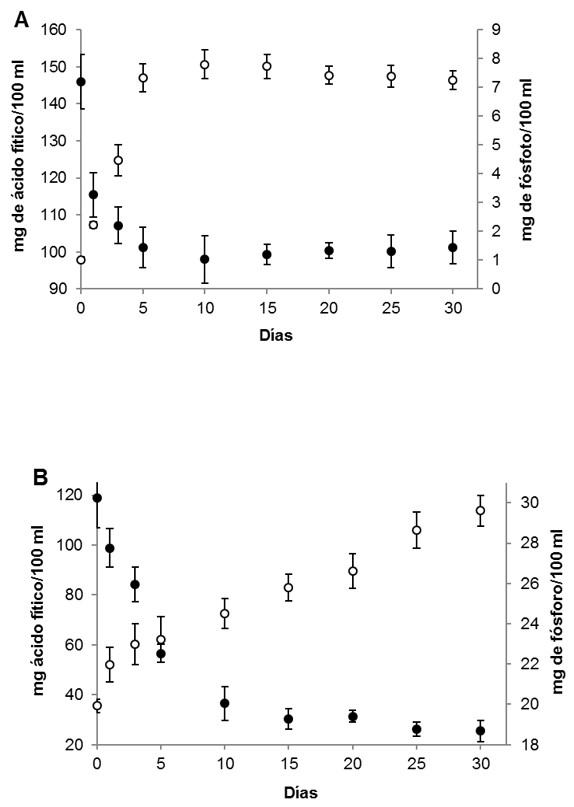
Figura 2. Evolución de la concentración de ácido fítico (●) y fósforo libre (○) durante la fermentación del repollo blanco (a) y del repollo chino (b).
Los diferentes comportamientos en la evolución de los parámetros estudiados se deben a las condiciones originadas durante la fermentación que influyen en la actividad de las fitasas vegetales y bacterianas. Las fitasas vegetales actúan principalmente durante la germinación con el objetivo de aprovechar la reserva de fósforo contenida en los fitatos y solo degradan 2 grupos fosfatos de los 6 que posee la molécula original 8.
Las fitasas de las BAL en cambio, son enzimas extracelulares y tienen un pH óptimo de actividad que varía entre 5,5 y 7,0. Valores de pH por debajo de 4,5 reducen sensiblemente la capacidad degradativa y a valores menores a 4,0 la actividad es casi nula 16,22,23. En los casos estudiados, el repollo blanco, como se señaló anteriormente, posee una mayor concentración de azúcares fermentables y en consecuencia mayor producción de ácidos orgánicos, en especial láctico. Luego del día 5, el pH continuó descendiendo registrándose en el día 10, un pH por debajo de 4,0. En forma paralela se puede observar que luego del día 5, no se registra un descenso de la concentración de ácido fítico ni un aumento en la de fósforo.
En el caso del repollo chino, el descenso del pH fue menos abrupto y no descendió a valores menores a 4,3 durante los 30 días de ensayo. El descenso marcado de la concentración de ácido fítico se produjo durante los primeros 10 días, sin embargo, el proceso degradativo continuó, en forma menos marcada, hasta el final de la experiencia. Como consecuencia de este fenómeno se observó que la concentración de fósforo aumentó en forma continua durante los 30 días.
En ambos casos se puede concluir que la actividad enzimática depende de las condiciones establecidas en la fermentación y del aumento poblacional de las BAL. La capacidad nutricional de los repollos permite una rápida duplicación celular y mayor oferta de fitasa extracelular, sin embargo, el metabolismo de carbohidratos trae como resultado un descenso marcado de pH y una menor actividad enzimática 14,16.
Determinación de hierro libre
En la Figura 3 se puede observar la evolución de la concentración de hierro libre en los vegetales estudiados durante la fermentación. Las concentraciones iniciales son comparables y estuvieron en valores menores a 0,05 mg/100 ml. Durante el desarrollo del proceso se observa un constante aumento de la concentración, sin embargo, el fenómeno es más marcado en el caso del repollo chino que alcanzó a los 30 días 0,29 mg/100 ml mientras que el repollo blanco exhibió una concentración de 0,16 mg/100 ml.
Estos distintos comportamientos tienen correlación con la variación de la concentración de ácido fítico, su degradación implica la liberación de hierro que, en forma de quelatos, resulta inaccesible para la absorción intestinal 11. El hierro liberado de la molécula de los fitatos tiene gran afinidad por el ácido ascórbico con el que forma quelatos estables y solubles facilitando su biodisponibilidad. Las brasicáceas contienen concentraciones de ácido ascórbico que no solo permiten esta función, sino que además su alto poder reductor transforma el hierro trivalente en bivalente, condición necesaria para ser transportado a través de la membrana del enterocito 12.
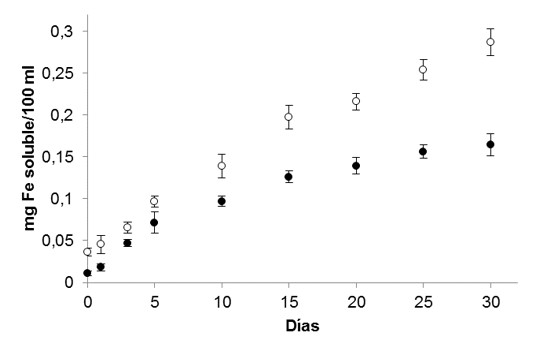
Figura 3. Evolución de la concentración de hierro soluble durante la fermentación del repollo blanco (●) y repollo chino (○).
CONCLUSIONS
Las fermentaciones en general y de las crucíferas en particular constituyen una forma fácil y económica para conservar vegetales por largo tiempo y en forma segura. Las BAL, el grupo más relevante en este proceso, no solo logran descender el pH a valores incompatibles para bacterias patógenas o deteriorantes sino que su metabolismo produce otros cambios favorables a la salud y/o nutrición. En este trabajo se determinó que, en un proceso de 30 días de fermentación, ambas brasicáceas exhibieron marcados descensos en las concentraciones de ácido fítico y, como consecuencia un aumento del fósforo soluble y de hierro libre. El efecto se observó en forma más marcada en el repollo chino, su menor oferta de carbohidratos no permite un descenso de pH tan abrupto como en el repollo blanco y, en consecuencia, se mantiene en valores compatibles con la actividad enzimática de las fitasas.
Contribuciones de los autores: Conceptualización, Parada R., Vallejo M. y Marguet E.; metodología, Marguet E.; software, Parada R..; validación, Parada R., Sosa F., Marguet E. y Vallejo M.; análisis formal, Parada R y Marguet E.; investigación, Parada R., Sosa F., Marguet E. y Vallejo M.; curación de datos, Parada R. y Marguet E.; redacción — preparación del borrador original, Parada R. y Marguet E..; redacción — revisión y edición, Parada R., Sosa F., Marguet E., Vallejo M.; visualización, Parada R., Sosa F. ; supervisión, Parada R, Marguet E., Vallejo M..; administración de proyectos, Vallejo M. y Marguet E..; adquisición de financiación, Vallejo M. y Marguet E. Todos los autores han leído y aceptado la versión publicada del manuscrito
Financiamiento: Este trabajo fue financiado por la Universidad Nacional de la Patagonia San Juan Bosco (SCT PI N°1519), por el Ministerio de Educación-Secretaría de Políticas Universitarias (UNPSJB7744) y por el Ministerio de Ciencia, Tecnología e Innovación (FONCyT PICT-2019-01348).
Conflictos de Interés: Los autores declaran no tener conflictos de interés.
REFERENCIAS
1. Di Cagno, R.; Coda, R., De Angelis, M.; Gobbetti, M. Exploitation of vegetables and fruits through lactic acid fermentation. Food Microbiol. 2013, 33, 1–10.
2. Settanni, L.; Corsetti, A. Application of bacteriocins in vegetable food biopreservation. Int. J. Food 2008, 121, 123–138.
3. Septembre-Malaterre, A.; Remize, F.; Poucheret P. Fruits and vegetables, as a source of nutritional compounds and phytochemicals: Changes in bioactive compounds during lactic fermentation. Food Res. Int. 2018, 104, 86–99.
4. Fernández-León, A.M.; Lozano, M.; González, D.; Ayuso, M.C.; Fernández-León, M. F. Bioactive compounds content and total antioxidant activity of two savoy cabbages. Czech. J. Food Sci. 2014, 32, 549–554.
5. Traka, M.H. Health Benefits of Glucosinolates. In: Glucosinolates, Kopriva SB (ed.); Academic Press, 2016, Volume 80, pp 247–279 .
6. Vale, A.P.; Santos, J.; Brito, N.V.; Fernandes, D.; Rosa, E.; Olivera, MB. Evaluating the impact of sprouting conditions on the glucosinolate content of Brassica oleracea sprouts. Phytochemistry 2015, 115, 252–260.
7. Gemede, F.H.; Ratta, N. Antinutritional Factors in Plant Foods: Potential Health Benefits and Adverse Effects. Int. J. Nutr. Food Sci. 2014, 3, 284–289.
8. Rao, D.; Rao, K.V.; Reddy, T.P.; Reddy, V.D. Molecular characterization, physicochemical properties, known and potential applications of phytases: An overview. Crit. Rev. Biotechnol. 2009, 29, 182–198.
9. Kumar, V.; Sinha, A.; Makkar, H.; De Boeck, G.; Becker, K. Phytate and phytase in fish nutrition. J. Anim. Physiol. Anim. Nutr. 2011, 96, 335–364.
10. Kumar, A.; Chanderman, A.; Makolomakwa, M.; Perumal, K.; Singh, S. Microbial production of phytases for combating environmental phosphate pollution and other diverse applications. Crit. Rev. Environ. Sci. Technol. 2016, 46, 556–591.
11. Kumar, V.; Sinha, A.K.; Makkar, H.P.S.; Becker, K. Dietary roles of phytate and phytase in human nutrition: A review. Food Chem. 2010, 120, 945–959.
12. Teucher, B.; Olivares, M.; Cori, H. Enhancers of iron absorption: ascorbic acid and other organic acids. J. Int. Vitaminol. Nutr. 2004, 74, 403–419.
13. Azeem, M. Microbial phytase activity and their role in organic P mineralization. Arch. Agron. Soil Sci. 2015, 61, 751–766.
14. Singh, B; Kunze, G; Satyanarayana, T. Developments in biochemical aspects and biotechnological applications of microbial phytases. Biotechnol. Mol. Biol. Rev. 2011, 63, 69–87.
15. Riad, G.; Ahmed, A.; El-Baky, M.; Ghoname ElSayed, A., Hegazi A. Cabbage Nutritional Quality as Influenced by Planting Density and Nitrogen Fertilization. Fruit. Veg. Cereal. Sci. Biotech. 2009, 3, 68-74.
16. Marguet, E.R.; Ledesma, P.; Vallejo, M. Disponibilidad de fósforo soluble en ensilado experimental fermentado con una cepa de Lactococcus lactis subsp. lactis con alta actividad de fitasa. Rev. Soc. Ven. Microb. 2013, 33, 116–121.
17. Mikulski, D.; Klosowski, G. Phytic acid concentration in selected raw materials and analysis of its hydrolysis rate with the use of microbial phytases during the mashing process. J. Inst. Brew. 2015, 121, 213–218.
18. Braunschweig, J.; Bosch, J.; Heister, K.; Kuebeck, C.; Meckenstock, R.U. Reevaluation of colorimetric iron determination methods commonly used in geomicrobiology. J. Microbiol. Methods 2012, 89, 41–48.
19. USDA. United States Department of Agriculture 2020. FoodData Center. In https://fdc.nal.usda.gov/fdc-app.html#/?query=cabbage. [Accessed 25 October 2020].
20. Christiansen, J.K.; Hughes, J.E.; Welker, D.L.; Rodríguez, B.T.; Steele, J.L.; Broadbent, J.R. Phenotypic and genotypic analysis of amino acid auxotrophy in Lactobacillus helveticus CNRZ 32. Appl. Environ. Microbiol. 2008, 74, 416–423.
21. Fossi, B.T.; Akwanwi, C.M.; Takop, N.G.; Bille, E.Y.; Wanji, S. Phytic acid degradation by selected lactobacilli isolated from fruits and vegetables and their potential as probiotics. Eur. J. Biotechnol. Biosci. 2016, 4,34–41.
22. Haros, M.; Bielecka, M.; Honke, J.; Sanz Y. Phytate-Degrading Activity in Lactic Acid Bacteria. Polish J. Food Nutr. Sci. 2008, 58,33–40.
23. De Angelis, M.; Gallo, G.; Corbo, M.R.; McSweeney, P.L.H.; Faccia, M.; Giovine, M. et al. Phytase activity in sourdough lactic acid bacteria: purification and characterization of a phytase from Lactobacillus sanfranciscensis CB1. Int. J. Food Microbiol. 2003, 87,259–270.
Received: 14 Agoust 2021 / Accepted: 27 March 2021 / Published:15 May 2022
Citation: Parada, R.; Sosa, F.; Marguet, E: Vallejo, M. Cambios en la concentración de ácido fítico, fósforo libre y hierro soluble durante la fermentación de repollo blanco y repollo chino. . Revis Bionatura 2022;7(2) 3. http://dx.doi.org/10.21931/RB/2022.07.02.3