CS 2019.02.01.29
Files > Conference Series > 2019 > Humboldt Kolleg 2019
Bionatura Conference Series Vol 2. No 1. 2019
“Breaking Paradigms: Towards a Multi-, Inter- and Transdisciplinary Science” In commemoration of the 250th Anniversary of Alexander von Humboldt
Population Estimates and Habitat Selection of White-tailed Deer (Odocoileus virginianus ustus) in the Antisana Water Conservation Area, Ecuador

*Markus Tellkamp1, Henry Herrera2, Ernesto Ponsot3, Isidro Bautista4
available in: http://dx.doi.org/10.21931/RB/CS/2019.02.01.29
ABSTRACT
The Antisana Water Conservation Area (ACHA) greatly contributes to the water supply of the Metropolitan District of Quito (DMQ). Therefore, integrated management of the area needs to take into account the ecological interactions that sustain the interception and slow release of water by the vegetation and soil. An apparent increase in the population of white-tailed deer (Odocoileus virginianus) has led to the concern that this herbivore may alter the vegetation and thus the water cycle. In this study, we obtain the first population estimates for white-tailed deer in ACHA and the second estimate for the Ecuadorean Andes. Vantage point counts (VPC) set the minimum population for the reserve at 760 individuals, whereas the fecal standing crop (FSC) indirect method determined an estimate of 953 ± 368 (90% confidence interval) for a density of 11.27 individuals/km2. The density of deer is low compared to other sites in the Colombian and Venezuelan Andes. From a management perspective, the optimal number of deer for the reserve will depend on the impact the deer have on the ideal composition of plants for the long-term production of water.
Keywords: White-tailed deer, Odocoileus virginiatus ustus, population estimates, vantage point counts, fecal standing crop
INTRODUCTION
The human population of the Metropolitan District of Quito (DMQ) is growing at a rapid rate, especially in the urban districts. Here, the demand of water is expected to grow by 28.5% between 2020 and 20401. In addition, a reduction in glaciers from nearby mountains due to climate change2, threatens the district’s ability to supply water for this increasing demand. The Fondo de Agua (FONAG) and the Empresa Pública Municipal de Agua Potable y Saneamiento (EPMAPS) have thus established three reserves for the conservation of water, totaling nearly 20,000 hectares. One of these areas, the Área de Conservación Hídrica Antisana (ACHA) has a capacity of 1650 l/s of water for the DMQ3. This area is part of the Reserva Ecológica Antisana and was formally part of a large hacienda, supporting tens of thousands of livestock4.
Because of the past presence of large numbers of sheep, cows, and horses, much of the ACHA consists of an unusual mix of habitats for the Ecuadorian Andes. Instead of large areas of Calamagrostis intermedia bunchgrass, the landscape is composed of extensive areas of cushion plants (locally referred to as almohadones) and herbaceous vegetation mixed in with a low Calamagrostris (cf. C. fibrovaginata) grass. The composition of the vegetation has a great impact on the water cycle of the páramo ecosystem. For example, a change from bushy vegetation to agricultural fields represents a reduction of up to 40% in water retention ability in lab dried soil and 21% in cultivated soils5. The proper revegetation of the ACHA is thus a priority for the FONAG4, whose staff is now actively removing the remaining individuals of livestock from the area. This removal, however, has important consequences for food chains based on carrion and has the potential to affect the populations of iconic species, such as the Andean Condor (Vultur gryphus) and the curiquingue or Carunculated Caracara (Phalcoboenus carunculatus). In addition, released from competition and predation pressures (predators were hunted to extirpation to protect the livestock), white-tailed deer are now making a comeback, spawning concerns that they may have a detrimental impact on the recovering páramo ecosystem. Although grazing at low levels appears to have little impact on water retention in páramos soils6, it is unclear what impact deer grazing may have at ACHA, especially after decades of intense livestock grazing.
Large herbivores particularly white-tailed deer (Odocoileus virginianus s.l.), can determine vegetation structure and composition, especially when overabundant7. Although little is known about the effects on grasslands or prairies, deer usually benefit unpalatable or rapidly growing species and reduce palatable and/or slowly growing species of plants. These effects have been observed with other species of deer as well. In grasslands of the Swiss Alps, Schütz et al.8 further found that red deer (Cervus elapus) grazing pressure tended to increase the diversity of plants. They found that most plants able to resist grazing were of low stature and were characterized by anti-herbivore protection mechanisms and/or short life spans. Previous research in forest ecosystems has shown that interactions between large herbivores and ecosystems are complex and not easily predictable. In North American rangelands, changes in grazing pressures have been shown to exhibit non-linear effects on vegetation: plant communities may reach multiple stable states and a decrease in grazing might not lead to the recuperation of the ecosystem to a former climax-like state9. Similar non-linear changes have been observed for other ecosystems as well10. For the páramo grasslands of Ecuador we still lack a basic understanding of the herbivore-ecosystem interactions, both for domestic and wild ungulates. In other words, we do not even have a basic understanding of how herbivores affect páramo vegetation.
At ACHA it has become a necessity to monitor the increasing population of white-tailed deer and its potential impact on the vegetation and thus the entire ecosystem. Deer have been a natural component of páramo grasslands for millions of years, but we do not fully understand its ecological role in this poorly understood ecosystem. There is still great disagreement over what a “natural” páramo grassland – without interference of humans – would look like11. Here we present the first, preliminary estimates for white-tailed deer at the ACHA and provide a framework for ascertaining the ideal population size for the area. It is our hope that we will be able to provide the FONAG with a fast, easy and reliable methodology to monitor the deer population at the ACHA over the long term.
Methods and Study Site
Estimates of deer population size can be obtained by direct and indirect methods. The former are based on visual deer counts and allows for estimation of sex ratios and age structure of the population. Estimates obtained by these methods are a snapshot of the population and the number of deer observed is valid only for the day of the survey12. We conducted Vantage Point Counts (VPCs) because in the open grasslands in mountainous terrain, deer can be seen at great distances. Indirect methods for deer are based on counts of signs or marks, usually fecal pellet groups. Estimates obtained by indirect means are usually an average representing the population size over several months12. Of the three indirect methods we are using in our study, Fecal Standing Crop (FSC), Fecal Accumulation Rate (FAR) and Distance methods, here we only present data for the FSC method.
Vantage Point Counts
Vantage Point Counts (VPCs) consist of observing animals from a point with clear vision of the surroundings13 and they are perfectly suited for the mountainous grasslands ACHA. VPCs were repeated four times, as recommended by Mayle et al.13 (Figure 1), in eight general areas within ACHA from dawn to 09:00 and 15:00 to dusk on 12 December 2018 and 13 December 2018. Observations lasted at least three hours to increase the likelihood that deer, which had lain down to ruminate before the onset of the count, would become active and thus visible during the count13. Each team carefully and silently scanned the surrounding area for the presence of deer with binoculars. Three groups were also equipped with spotting scopes. Information recorded included deer activity, number of individuals, sex, age, habitat and weather. All groups were provided with medium range two-way radios to report on deer movements and thus avoid double counting deer coming from other areas.
We selected ten vantage points (Figure 1) in areas known for having high deer densities according to the guardaparamos of FONAG, EPMAPS, and the Ministry of Environment (MAE). The main criterion for the selection was to find the largest number of deer possible within the 8457 ha reserve. We did not choose locations randomly as the logistics of placing a large number of observers in the field are challenging and expensive14,15. Also, high-altitude environments with low levels of oxygen pose additional constraints on intense sampling within a large area16. Each vantage point covered an area within a radius of approximately one kilometer; however, we are still estimating the effective area surveyed in this census.
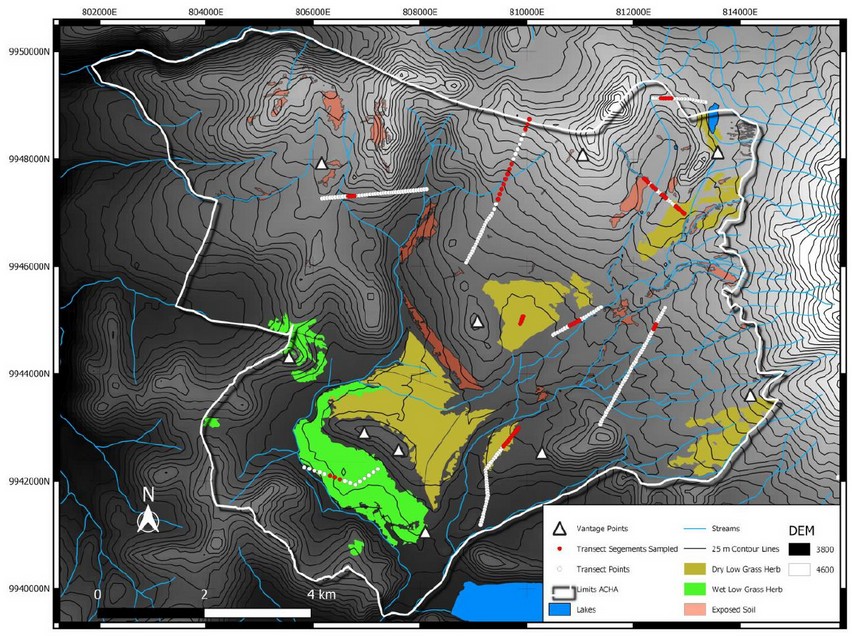
Figure 1. Map showing the limits of the Área de Conservación Hídrica Antisana (ACHA), the location of the vantage count points and the sampled segments (red) of the transects (unsampled segments indicated by white dots). Digital Elevation Model is shown in shades of gray, with darker shades indicating lower elevations. Contour lines are at 25-meter intervals.
Indirect Counts Using Pellet Groups
Nine transects, totaling 14,150 meters, were placed and oriented systematically within ACHA as recommended by Marques et al. 12 covering the majority of the altitudinal gradient, the most representative habitats, and areas with known high deer densities, making sure we were covering areas of low density as well. The transects were marked every 50 or 100 meters using PVC tubes. During counts, the centerline of the transect was marked with a rope and the distance to either side measured with measuring tapes. The selected width for the transects was 4 meters, 2 meters on either side of the transect. Two or three observers searched for pellet groups.
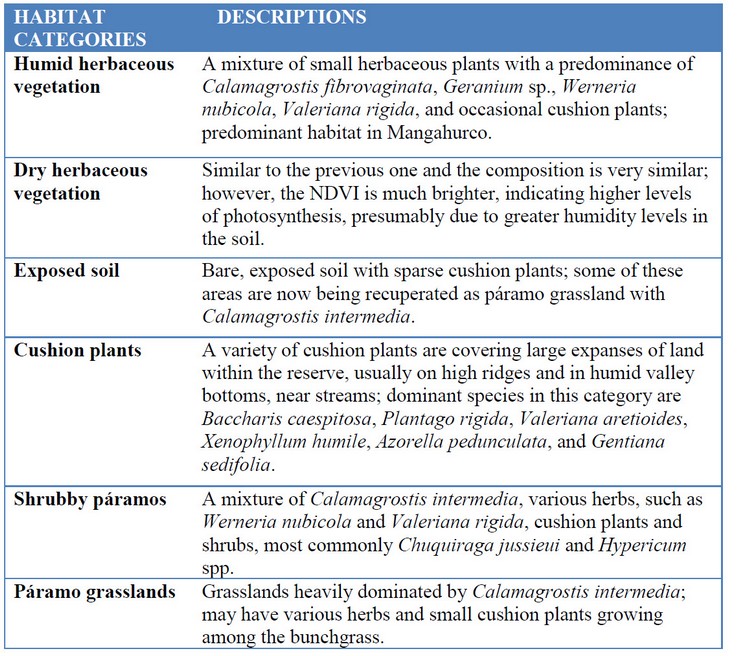
Within the 8457 ha area of ACHA we recognize six habitats that we consider important for the use and distribution of deer within the reserve: humid herbaceous vegetation, dry herbaceous vegetation, exposed soil, cushion plants, shrubby páramos and páramo grasslands (Table 1). We are in the process of digitizing polygons for these habitats to estimate the size of the area covered by each of them; however, as of now we only have area estimates for humid herbaceous vegetation, dry herbaceous vegetation, and exposed soil (Table 2). In this study, we pool data for the remaining habitats, which we have similar pellet densities. Habitats were classified on the basis of RapidEye satellite imagery, particularly choices calculation of the Normalized Difference Vegetation Index (NDVI) in QGIS software. We do not use the habitat classification by the FONAG because, based on our observation, it does not reflect the deer’s’ habitat choices accurately.
Fecal Standing Crop (FSC)
FSC is based on counts of all accumulated pellets groups within sampling plots17 and assumes that all objects are seen and recorded18. Our analyses presented here are based on surveys of 24.5 sampling plots (each complete plot measures 100 m x 4 m), for a total sample area of 9400 m2 in four habitat categories (Table 2). Only fecal pellet groups consisting of six or more pellets were considered17, and those lying on the edges of the sampling plots were recorded when the center of the group was inside the plot. Fecal pellet group density was calculated as follows:





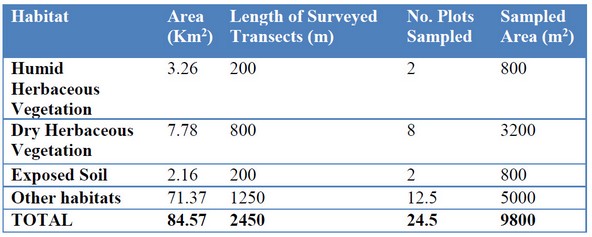
Table 2. Details of the transects surveyed using FSC through counts of pellet groups
RESULTS
Vantage Point Counts
Detectability of animals may vary due to weather conditions, nearby agricultural practices, the experience of the observers, and types of vegetation15,16. The first day at dawn, for example, fog and rain at most vantage points decreased deer detectability. In contrast, during a mostly sunny afternoon on the second-day observers detected more deer. Mayle et al. 13 recommend using the maximum value counted as the final estimate; therefore, we consider the 760 deer seen on 13 December 2018 during the afternoon to be the minimum abundance estimation at ACHA (Table 3). The vantage point at Mangahurco yielded the highest number of sightings with 259 deer, followed by Antena with 225 deer. With the exception of Laguna Santa Lucía, where only 22 deer were counted, the remaining vantage points produced 40-60 deer observations. These numbers provide us an idea of the deer’s habitat preference within ACHA. Sites dominated by humid herbaceous vegetation supported the most deer, whereas sites with lower counts were composed of a mix of cushion plants, C. intermedia páramo grass, and shrubs.
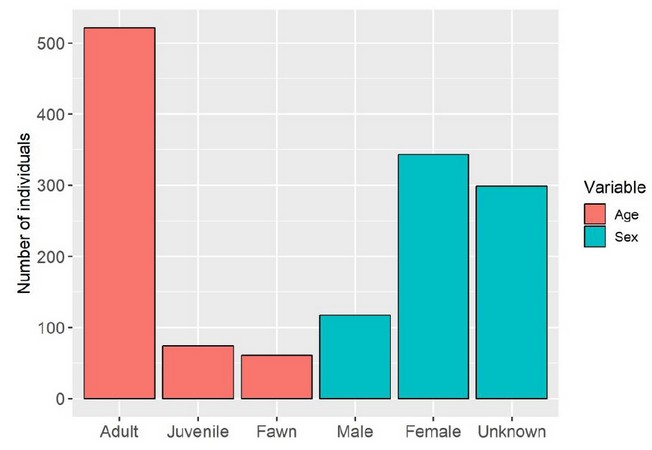
Figure 2. Overall age structure and sex distribution of white-tailed deer at ACHA.
Moreover, this method gives us insights on sex ratios and age structure. The number of females outnumber males about 3 to 1. Similarly adults outnumber juveniles and fawns 6 to 1 (Figure 2). The vantage point Antena stands out because of an unusual high number of female deer and fawns. Perhaps this area presents the best condition in the ACHA for females to nurse their young (Table 4).
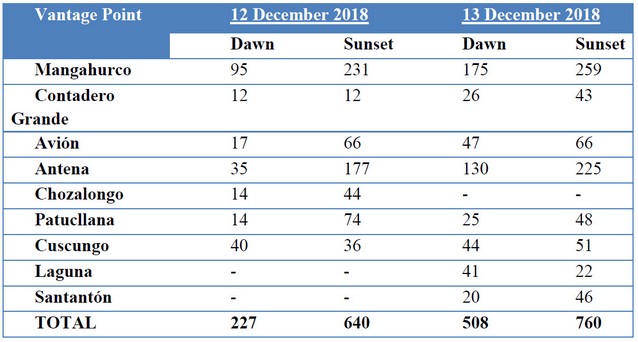
Table 3. Number of white-tailed deer counted at the different sites during the vantage point counts.
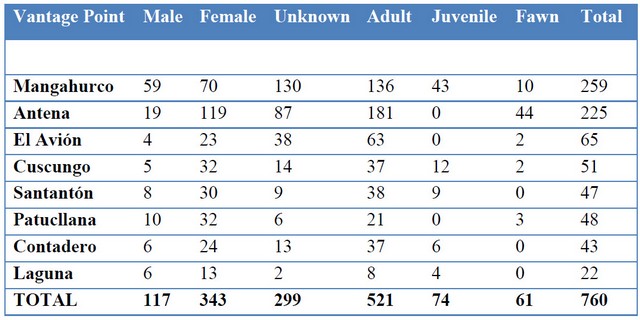
Table 4. Sex and age groups of deer by vantage point (see Figure 1).
Fecal Standing Crop
Our preliminary estimate for the average length time for pellets to decay at ACHA as of March 2019 is 240 days. This estimate is just a little bit lower than the prospective value (277.80 days) estimated by Mateus21 at PNNC. However, this value may change upward as so-called prospective estimates can vary due to factors such as rainfall, diet, among others20. The habitat with the highest pellet groups density was humid herbaceous vegetation has the highest pellet group density; however, its area only represents 3.9% of the entire area so it does not have the highest absolute deer abundance (307 individuals; Table 5). This value is surprisingly similar to the number estimated during the visual census (305 individuals) at the same vantage points (Mangahurco and Santantón), which include most of the humid herbaceous vegetation at ACHA.
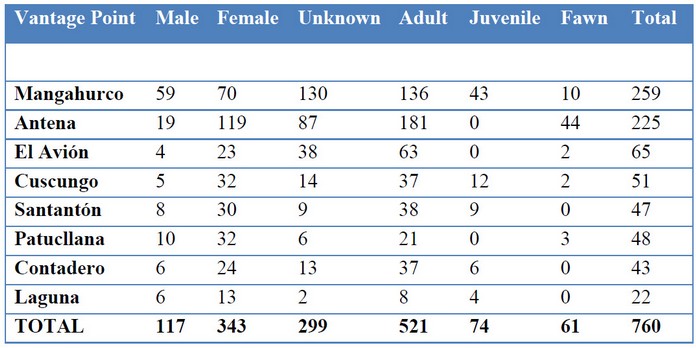
Table 5. Pellet numbers and densities and their corresponding estimates for deer densities and abundances in different habitats using FSC.
The dry herbaceous vegetation habitat has the second-highest deer density (43.38 individuals/km2). This information suggests that deer prefer habitats with low herbaceous vegetation. In contrast, habitats composed of páramo grasslands, cushion plants, and shrubby grasslands have low deer densities (4.26 individuals/km2). Because their summed areas represent 84% of the entire study area, these habitats harbor a considerable number of deer (304). Finally, exposed soils have the lowest deer density (2.24 individuals/km2). Finally, our preliminary deer abundance estimation at ACHA using the FSC method with a 90% of confidence is 953 ± 368 and the corresponding density is 11.27 individuals/km2. We hope that the precision of this estimate will increase as more sampling plots are surveyed and the areas of each sampled habitat are better defined, so the results will be shown in the future.
DISCUSSION
Being only the second attempt to gauge the size of deer populations in the Ecuadorean páramo, this study provides the first estimates of the deer population in ACHA. Direct, simultaneous observations suggest the minimum population is 760 individuals. Indirect measures suggest a population of close to 1000 individuals, which corresponds to a population density of 11.27 individuals/km2. These numbers are considerably higher than those reported by Albuja22, but are not unusual for sites in the Colombian and Venezuelan Andes23.
Even though our study strongly suggest the humid herbaceous vegetation is the preferred habitat, density may not always be a good indicator of habitat quality or preference24. In this study, we hypothesize that high densities of white-tailed deer at ACHA correlate with habitat quality and preference. The humid herbaceous vegetation has a high proportion of forbs, a group of plants that is preferred by white-tailed deer elsewhere25. At ACHA, these plans also appear to be highly productive as judged by the Normalized-Difference Vegetation Index (data not shown). This vegetation is also easily accessible by wide-ranging deer. In contrast, the tall C. intermedia grass does not provide much fresh forage and is difficult to move through, cushion plants are often spikey and have tough leaves, brushy páramo consists largely of C. intermedia and bushes with spikey and tough leaves (such as Chuquiraga jussieui), and exposed soil contains few plants. Tall C. intermedia grass is, however, an important feature for deer near the herbaceous vegetation as it appears to be a good place for deer to hide from predators and take shelter from the bad weather. Another caveat is that the results of direct counts, which are limited to two a few days, may also provide misleading information about habitat use12; however, results from FSC suggest that our observations are representative for ACHA over longer periods.
The ACHA was created to safeguard water production for the city of Quito. Thus, the area needs to be managed to maximize this ecosystem service. The question arises, what is the optimal population of deer that allows managers to meet this goal? At this point, it is difficult to ascertain the impact of deer on the ACHA ecosystem. The population is not unusually high, within the bounds of what has not raised any concerns in Colombia and Venezuela. From a management perspective, the ideal number of deer at the reserve depends on what habitats FONAG and EPMAPS want to have represented at ACHA. This, in turn, will depend on the ideal composition of vegetation based on the capacity of different vegetation types to capture and slowly release water. For instance, cushion plants have been found to increase the transition time of water26. It might therefore not be desirable to convert the large expanses of cushion plants to grasslands. We do not yet understand the role of large herbivores in creating and maintaining these (in our experience) unusually large expanses of cushion plants at ACHA. Are deer important in maintaining them? Cottontails, which undergo large populational cycles in this páramo, seem to be detrimental to them (Francisco Black and Manuel Simba, personal comments). Deer seem to favor the valley bottoms, characterized by herbaceous vegetation and C. fibrovaginata or little bunches of C. intermedia. These areas also appear to be favored by Black-faced Ibises (Theresticus melanopis) and Curiquingues (Phalcoboenus carunculatus), two species of some conservation concern. Would the vegetation in these valleys be the same without the deer or low densities of deer? It is our hope that with the protocols we develop in this study to monitor deer populations at ACHA will help the FONAG and EPMAPS to answer some of these pressing questions in the long term.
CONCLUSION
The density of 11.27 individuals/km2 deer at ACHA is still low compared to other sites in the Andes and elsewhere, although locally as in in the humid herbaceous vegetation the density can be very high (94.05 individuals/km2). The bulk of the reserve is home to as many deer as the two herbaceous types of habitat, respectively (Table 5). The vegetation with the highest densities is the most productive one as judged by NDVI and probably consists of mostly fast growing species, highly resistant to grazing. Thus, presently deer do not pose a threat to the vegetation. Future increases, however, might be highly detrimental7. Ultimately, the number of deer that the reserve should hold depend on the management goals of FONAG and EPMAPS. These goals will depend on what combination of vegetation types favors the greatest levels of water capture and slowest rates of water release, and how deer contribute to maintaining them in the long term.
REFERENCES
1. EPMAPS. Estudios de actualización del plan maestro integrado de agua potable y alcantarillado para el DMQ - Resumen ejecutivo. Quito: EPMAPS; 2011. 61 p.
2. Vuille M, Francou B, Wagnon P, Juend I, Kaser G, Mark BG, Bradley RS. Climate change and tropical Andean glaciers: Past, present and future. Earth Sci Rev. 2008; 89: 79–96.
3. EPMAPS. Captación [Internet]. Quito: EPMAPS; 2018. [cited 2019 May 21]. Available from https://www.aguaquito.gob.ec/captacion/.
4. Aguirre N, Torres J, Velasco P. Guía para la Restauración Ecológica en los Páramos del Antisana. Quito: Fondo para la Protección del Agua-FONAG; 2014. 64 p.
5. Buytaert W, Deckers J, Dercon G, De Bièvre B, Poesen J, Govers G. Impact of land use changes on the hydrological properties of volcanic ash soils in South Ecuador. Soil Use Manag. 2002; 18: 94–100.
6. Crespo P, Célleri R, Buytaert W, Feyen J, Iñiguez V, Borja P, De Bièvre B. Land use change impacts on the hydrology of wet Andean páramo ecosystems. In: Holko L, Schumann SA , Littlewood I, Pfister L, Warmerdam P, Schroder U, Herrmann A, editors. Status and Perspectives of Hydrology in Small Basins. Proceedings of the Workshop; Goslar-Hahnenklee, Germany; 2009 March 30–April 2. London: International Association of Hydrological Sciences; 2010. IAHS Publ. 336: 71–76.
7. Côté, SD. Impacts on ecosystems. In: Hewitt DG, editor. Biology and management of white-tailed deer. Boca Raton: CRC Press; 2011. p. 379–398.
8. Schütz M, Risch AC, Leutzinger E, Krüsi BO, Achermann G. Impact of herbivory by red deer (Cervus elaphus L.) on patterns and processes in subalpine grasslands in the Swiss National Park. For Ecol Manage. 2003; 181: 177–-188.
9. Laylock WA. Stable states and thresholds of range condition on North American rangelands: A viewpoint. J Range Manage. 1991; 44: 427–433.
10. Côté SD, Rooney TP, Tremblay J-P., Dussault C, Waller DM. Ecological impacts of deer overabundance. Annu Rev Ecol Syst. 2004; 35: 113–147.
11. Gade DW. Nature and culture in the Andes. Madison: University of Wisconsin Press; 1999. p. 302.
12. Marques FFC, Buckland ST, Goffin D, Dixon CE, Borchers DL, Mayle BA, et al. Estimating deer abundance from line transect surveys of dung: sika deer in southern Scotland. J Appl Ecol. 2001; 38: 349–63.
13. Mayle BA, Peace AJ, Gill RMA. How many deer? A field guide to estimating deer population size. Edinburgh: Forestry Commission; 1999. 96 p. (Field Book series; no.: 18).
14. Nichols JD, Hines JE, Sauer JR, Fallon FW, Fallon JE, Heglund PJ. A double-observer approach for estimating detection probability and abundance from point counts. Auk Ornithol Adv. 2000; 117: 393–408.
15. Zaccaroni M, Dell’Agnello F, Ponti G, Riga F, Vescovini C, Fattorini L. Vantage point counts and monitoring roe deer: Monitoring Roe Deer. J Wildl Manag. 2018; 82: 354–61.
16. Singh NJ, Milner-Gulland EJ. Monitoring ungulates in Central Asia: current constraints and future potential. Oryx. 2011; 45: 38–49.
17. Smart JCR, Ward AI, White PCL. Monitoring woodland deer populations in the UK: an imprecise science. Mammal Rev. 2004; 34: 99–114.
18. Burnham KP, Anderson DR. The Need for Distance Data in Transect Counts. J Wildl Manag. 1984;48: 1248–54.
19. R Core Team. R: A language and environment for statistical computing [Internet]. Vienna, Austria: R Foundation for Statistical Computing; 2017. [cited 2019 July 25]. Available from: https://www.R-project.org/
20. Laing SE, Buckland ST, Burn RW, Lambie D, Amphlett A. Dung and nest surveys: estimating decay rates. J Appl Ecol. 2003; 40: 1102–11.
21. Mateus Gutiérrez C. Efecto de la estructura del hábitat sobre las características demográficas de dos poblaciones locales de Venado Cola Blanca, Odocoileus virginianus goudotii, en el Parque Nacional Natural Chingaza (Colombia) [master’s thesis]. Bogotá: Universidad Nacional de Colombia; 2014. p. 127.
22. Albuja LV. Biología y ecología del venado de cola blanca (Odocoileus virginianus ustus Gray, 1874) en los páramos de Oyacachi-Papallacta y Antisana, Ecuador. Politécnica. 2007; 27 Biología 7: 34-57.
23. Ortega-S JA, Mandujano S, Villarreal-González JG, Di Mare MI, López-Arevalo H, Molina M, Correa-Viana M. Managing white-tailed deer: Latin America In: Hewitt DG, editor. Biology and management of white-tailed deer. Boca Raton: CRC Press; 2011. p. 565–597.
24. Van Horne B. Density as a misleading indicator of habitat quality. J. Wildl. Manag, 1983; 47: 893–901.
25. Kie JG, Drawe DL, Scott G. Changes in diet and nutrition with increased herd size in Texas white-tailed deer. J Range Manage. 1980; 33: 28–34.
26. Mosquera GM, Segura C, Vaché KB, Windhorst D, Breuer L, Crespo P. Insights into the water mean transit time in a high-elevation. Hydrol Earth Syst Sci. 2016; 20, 2987–3004.
Received: 1 April 2019
Accepted: 28 May 2019
*Markus Tellkamp1, Henry Herrera2, Ernesto Ponsot3, Isidro Bautista4
1. Doctor en Zoología. Escuela de Ciencias Biológicas e Ingeniería. Carrera de Biología. Universidad Yachay Tech. [email protected]
2. Estudiante de pregrado de Biología. Escuela de Ciencias Biológicas e Ingeniería. Carrera de Biología. Universidad Yachay Tech. [email protected]
3. Doctor en Estadística. Escuela de Ciencias Matemáticas y Computacionales. Carrera de Matemáticas. Universidad Yachay Tech. [email protected]
4. Guardapáramo, Área de Conservación Hídrica Antisana. Fondo Para la Protección del Agua (FONAG).
*Corresponding author: Markus P. Tellkamp