2017.02.01.9
Files > Volume 2 > Vol 2 No 1 2017 > Revisiones
ARTÍCULO DE REVISIÓN
The Impact of Endocrine Disrupting Chemicals on the Environmental and their Potential Biotransformation by White-rot Fungi and their Oxidative Enzymes
El impacto de los compuestos disruptores endocrinos sobre el medio ambiente y su potencial biotransformación por hongos de pudrición blanca y sus enzimas oxidativas

Roberto Taboada-Puig1*, Maria Teresa Moreira2, Spiros N. Agathos1, Juan M. Lema2
Available from: http://dx.doi.org/10.21931/RB/2017.02.01.9
_______________________________________________________________________________________________________________________ ABSTRACT
The search of new technologies suitable for the treatment of wastewater containing endocrine disrupting chemicals (EDCs) such as bisphenol A (BPA), triclosan (TCS), estrone (E1), 17β-estradiol (E2) and 17α-ethinylestradiol (EE2) is a challenge since existing traditional wastewater treatment plants (WWTPs) are not able to eliminate them completely. Even at concentrations of ng/L, EDCs have an impact in the endocrine system of the fauna producing morphological deformities, reduced overall growth, reduced sperm quality and delayed ovulation, sex reversal male to female, among others. Efforts have been put in the study of different post-treatments to remove the residual concentration of EDCs present in WWTP effluents. The most frequently used technologies are advanced oxidation processes (AOPs) such as photocatalysis, photodegradation, sonolysis, ozonolysis, amongst others. In general, these processes have high degradation rate but they have low selectivity and high costs. In addition, AOPs can be a cause of concern itself since they may render harmful by-products or transformation products which can have similar or increased estrogenicity of that of the parent compound. A biological alternative may be use of white-rot fungi (WRF) or their lignin modifying enzymes (LMEs) to treat wastewater containing EDCs. From an operational point of view, the use of LMEs in in vitro systems, compared to the use of WRF in in vivo systems, is easier and cheaper since no aseptic conditions are needed. Among the LMEs, laccase is the most extensively studied enzyme for the degradation of BPA, TCS, E1, E2 and EE2 whereas peroxidases (LiP, MnP and VP) have not been studied in such level of detail.
Keywords: endocrine disrupting chemicals; white-rot fungi; lignin modifying enzymes; estrogenic activity.
_______________________________________________________________________________________________________________________
RESUMEN
La búsqueda de nuevas tecnologías adecuadas para el tratamiento de aguas residuales que contienen compuestos disruptores endocrinos (CDE), tales como bisfenol A (BPA), triclosán (TCS), estrona (E1), 17β-estradiol (E2) y 17α-etinilestradiol (EE2) es un reto, ya que las plantas tradicionales de tratamiento de aguas residuales tradicionales (EDAR) no son capaces de eliminarlas completamente. Incluso en concentraciones de ng/L, los CDEs tienen un impacto en el sistema endocrino de los animales produciendo deformidades morfológicas, reducción del crecimiento general, reducción de la calidad espermática y retraso de la ovulación, inversión sexual masculina a femenina, entre otros. Los esfuerzos se han enfocado en el estudio de diferentes post-tratamientos para eliminar la concentración residual de los CDEs presentes en los efluentes de la EDAR. Las tecnologías más empleadas son los denominados procesos de oxidación avanzada (POAs) tales como fotocatálisis, fotodegradación, sonolisis, ozonolisis, entre otros. En general, estos procesos tienen una alta tasa de eliminación pero tienen baja selectividad y alto coste. Además, los POAs pueden producir subproductos dañinos o productos con una estrogenicidad similar o mayor que la del compuesto inicial. Una alternativa biológica puede ser el uso de hongos de podredumbre blanca (HPB) o sus enzimas modificadoras de lignina (EML) para tratar aguas residuales que contienen CDEs. Desde un punto de vista operativo, el uso de EMLs en sistemas in vitro, en comparación con el uso de HPBs en sistemas in vivo, es más fácil y más barato ya que no se necesitan condiciones asépticas. Entre las EMLs, la lacasa es la enzima más ampliamente estudiada para la degradación de BPA, TCS, E1, E2 y EE2, mientras que las peroxidasas (LiP, MnP y VP) no se han estudiado en tanto detalle.
Palabras clave: compuestos disruptores endocrinos; hogos de podredumbre blanca; enzimas modificadoras de lignina; actividad estrogénica.
_______________________________________________________________________________________________________________________ INTRODUCTION
Endocrine disrupting chemicals
Since the end of the 1990 there has been a growing concern about the exposure to substances which are suspected to interfere with the endocrine system, and thus, may cause health effects such as cancer, behavioural changes and reproductive abnormalities in human beings and wildlife.
Many of the thousands of anthropogenic chemicals currently released into the environment are endocrine disrupting compounds (EDCs). These are defined as “a group of chemicals (natural, synthetic, industrial chemicals or by-products) present in the environment and suspected to alter the functions of the endocrine system and, consequently, causing adverse health effects in an intact organism, or its offspring or (sub) population”1. Figure 1 shows the main distribution of EDCs in the environment.
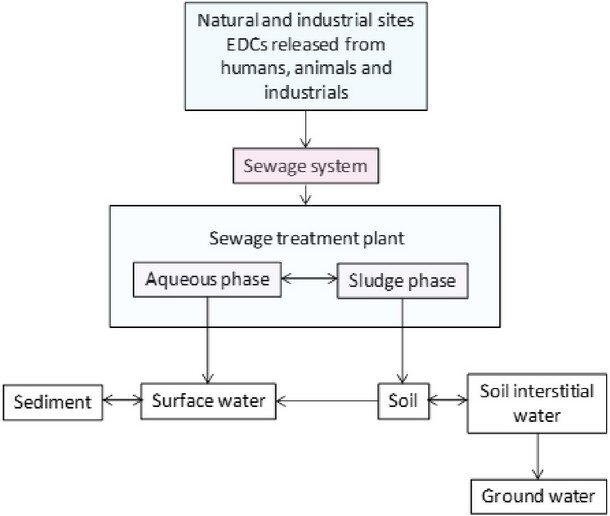
Figure 1. Distribution of EDCs in the environment.
Two classes of substances can cause endocrine disruption:
- Man-made substances which comprise: (i) synthetically-produced hormones, including oral contraceptives, such as ethynylestradiol (EE2), hormone replacement therapy and some animal feed additives, designed deliberately to interfere with and modulate the endocrine system; (ii) a variety of man-made chemicals, such triclosan (TCS) present in cleaning agents or bisphenol A (BPA) used in consumer goods and iii) various by-products from industrial processes such as dioxins.
- Natural hormones which include estrogens such as estrone (E1), 17β-estradiol (E2), and estriol (E3), progesterone and testosterone naturally found in the body of humans and animals, and phytoestrogens, such as isoflavonoides and coumestrol, present in some plants.
Endocrine systems regulate a multitude of developmental, metabolic, and reproductive processes including embryonic development, gonadal formation, sex differentiation, growth and digestion; therefore, endocrine disrupting compounds, may affect these processes by either binding to or blocking hormone receptors, thereby, triggering or preventing hormonal response.
Bisphenol A
Bisphenol A (BPA) is an industrial organic compound of formula (CH3)2C(C6H4OH)2 with two phenol functional groups (Figure 2).
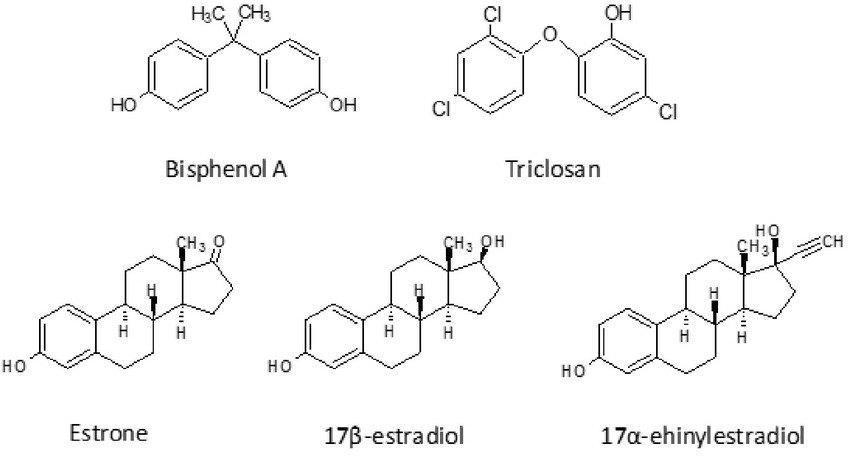
Figure 2. Molecular structure of bisphenol A, triclosan, estrone, 17β-estradiol and 17α-ethinylestradiol
Most BPA is used as an intermediate in the production of polycarbonate and epoxy resins, flame retardants, and other specialty products. Final products include adhesives, protective coatings, powder paints, automotive lenses, protective window glazing, etc. Polycarbonate is used in a number of household containers, including baby bottles, sippy cups, re-useable water bottles (sports bottles), pitchers, water carboys, tableware and food storage containers, whereas the epoxy resins act as a protective lining on the inside of metal-based food and beverage cans.
Health effects
The production and use of BPA is a controversial issue all around the world. The National Toxicology Program Center for the Evaluation of Risks to Human Reproduction (NTP) from the United States completed a review of BPA. Regarding BPA, the NTP expressed “some concern for effects on the brain, behavior, and prostate gland in fetuses, infants and children at current human exposures to bisphenol A”. The Program also expressed “minimal concern for effects on the mammary gland and an earlier age for puberty for females in fetuses, infant, and children at current human exposures to bisphenol A” and “negligible concern” for other outcomes. However, the Program stated that “additional research is needed to fully assess the functional, long-term impacts of exposures to bisphenol A on the developing brain and behavior”2.
In March 2012, the U.S. Food and Drug Administration (FDA) concluded that “the scientific evidence now does not suggest that the very low levels of human exposure to BPA through the diet are unsafe”. However, the FDA recognizes potential uncertainties in the overall interpretation of these studies; therefore, in July 2012, the FDA banned the use of BPA in baby bottles and sippy cups.
Canada was the first country to take action on BPA, thanks to the Chemicals Management Plan, developed by Canadian Environmental Protection Act (CEPA). Although there is no general recommendation concerning BPA, the Canadian Government advised to reduce exposure of newborns and infants less than 18 months3. Consequently, the proposed ban applies only to baby bottles made of polycarbonate. All other containers made with other types of plastics can continue to be used safely.
Concerning the use of bisphenol A in Europe, in 2006 the European Food Safety Authority (EFSA) concluded that infants aged 3 and 6 months fed using polycarbonate infant feeding bottles have the highest exposure to BPA, though below the tolerable daily intake (TDI, 0.05 mgBPA/kg). Even if the infant has sufficient capacity to remove BPA at worst-case exposure, the EFSA pointed out that an infant’s system to remove BPA is not as developed as that of an adult and it gradually reaches the adult capacity after 6 months. Consequently, the new EU regulation applied from May 1st 2011 prohibits the use of Bisphenol A in the manufacture or import of polycarbonate infant feeding bottles.
As a general conclusion, it can be established that even when it is not clearly demonstrated that BPA can have an effect on human beings, most countries around the world ban the use of BPA for the production of baby bottles or other household items that can be in close contact with infants.
Environmental risks
BPA emissions to the environment may be from several sources such as producing factories, installations that include BPA into plastic and leachates from plastic wastes and landfill sites. Variable ranges of BPA concentrations have been detected: 5–320 ng/L in river waters, 20–700 ng/L in sewage effluents, 2–208 ng/m3 in air, 0.2–199 ng/g in dust and 0.1–384 ng/g in food-stuffs. Municipal wastewater treatment helps achieve removal efficiencies in the range of 90-99%, depending on the technical capabilities of sewage treatment systems4.
Based on reported EC50 and LC50 values that range from 1.0 to 10 mg/L, BPA is classified as “moderately toxic” and “toxic” to aquatic biota by the European Commission and the United States Environmental Protection Agency (US EPA), respectively5. However, studies of BPA effects on wildlife indicate that the compound may be harmful even at environmentally relevant concentrations (12 μg/L) or lower6.
Triclosan
Triclosan is a type of bisphenol that exhibits antimicrobial activity (Figure 2). It is a synthetic, non-ionic, broad-spectrum antimicrobial agent, possessing mostly antibacterial, but also antifungal and antiviral properties7. Triclosan is fairly insoluble in aqueous solutions except for alkaline pH, and it is readily soluble in most organic solvents.
It is used in many contemporary and professional health care products, such as hand soaps, hand washing solution, deodorants, etc7. It is also included into fabrics and plastics, as those in toys, toothbrush handles, cutting boards, pizza-cutter and mop handles, as well as surgical drapes and hospital over-the-bed table tops.
Health effects
According to the FDA, there is no evidence that triclosan is hazardous to humans; there is no sufficient safety evidence to recommend changing consumer use of products that contain triclosan.
The Government of Canada completed its preliminary assessment of triclosan under the Canadian Environmental Protection Act and the Pest Controls Products Act. The review concludes that triclosan is not harmful to human health, but in significant amounts it may cause harm to the environment8.
The opinion of the European Commission, through the Scientific Committee on Consumer Products (SCPP), is that “taking into account the provided toxicological data, the continued use of triclosan as a preservative at the limit concentration of 0.3% in all cosmetic products is not safe for the consumer because of the magnitude of the aggregate exposure. However, its use at a maximum concentration of 0.3% in face products, toothpastes, hand soaps, body soaps/shower gels and deodorant sticks, is considered safe. However, the use of triclosan in other leave-on products (e.g. body lotions) and in mouthwash solutions is not considered safe for the consumer due to the higher level of exposure.”
Environmental risks
The widespread use of triclosan results in the discharge of this compound to wastewater. Triclosan is transported through the domestic waste stream to wastewater treatment plants (WWTPs9). Municipal wastewater treatment helps achieve removal efficiencies in the range of 51-95%, depending on the technical capabilities of sewage treatment systems10,11. Both the incomplete removal of triclosan from wastewater treatment plants and its presence in biosolids spread as fertilizers lead to triclosan being distributed in soils and surface waters.
Mass balance studies have demonstrated that triclosan also exhibits significant persistence, partitioning and sequestration in biosolids (logKOW~4.212), therefore 50±19% of the incoming triclosan, approximately, was observed to persist and become sequestered in biosolids in a typical WWTP comprising activated sludge treatment and anaerobic digestion13.
Natural and synthetic hormone substances: estrone, 17β-estradiol and 17α-ethinylestradiol
Estrogens are a group of steroid compounds, named for their importance in the oestrus cycle, functioning as the primary female sex hormone. While estrogens are present in both men and women, they are usually presented at significant higher levels in women of reproductive age14. The natural estrogens, estrone (E1) and 17β-estradiol (E2), and the synthetic one, 17α-ethinylestradiol (EE2), Figure 2, are the estrogens most commonly found in wastewater15.
Estrone, C18H22O2, also known as oestrone (3-hydroxy-1,3,5(10)-estretrien-17-one) is a C-18 natural steroid hormone. Estrone is one of the naturally occurring estrogens, the others being estradiol and estriol. Estrone is produced primarily from androstenedione originating from the gonads or the adrenal cortex and from estradiol by 17-hydroxysteroid dehydrogenase. 17β-estradiol, C18H24O2, also known as estradiol and oestradiol, (17β)-estra-1,3,5(10)-triene-3,17-diol, is another natural steroid hormone. Like other steroids, E2 is derived from cholesterol. 17α-ethinylestradiol, C20H24O2, is the major exogenous estrogen in humans. It is a bioactive estrogen used in several formulations of combined oral contraconceptive pills. It was approved by the FDA in the United States on 1943 and the FDA withdrew approval on 2004.
All humans as well as animals can excrete hormone steroids from their bodies, which end up in the environment through sewage discharge and animal waste disposal. Based on daily excretion of estrogen, dilution factor and previous measurements, levels of estrogens in ng/l are expected to be present in aqueous environmental samples14. Those steroids have been detected in influents and effluents of sewage treatment plants and surface water; therefore, since the three estrogens are not completely removed by WWTPs they remain with fluctuating concentrations in effluent, and discharge of such effluent. This may be the main reason for the wide occurrence of estrogens in surface waters, ground waters and even in drinking waters16.
White-rot fungi
White-rot fungi (WRF) belong to the class of basidiomycetes and certain ascomycetes, and they constitute the most important rotting fungi since they are the only microorganims able to mineralize lignin producing carbon dioxide and water. The term “white-rot” has been traditionally used to describe forms of wood decay, leaving a light, white, rather fibrous residue completely different from the brown powder left by brown rot fungi. Generally, WRF are unable to use lignin as a sole carbon source but they degrade it in order to gain access to cellulose and hemicellulose. Within this group, Phanerochaete chrysosporium is the most extensively studied species, although other fungi such as Bjerkandera adusta, Trametes versicolor, Pleurotus ostreatus are also well-known17.
Delignification is based on the WRF capacity to produce one or more extracellular lignin-modifying enzymes (LMEs) which, thanks to their lack of substrate specificity, are also capable of degrading a wide range of xenobiotics also at relatively low concentrations since they are not induced by either lignin or other related compounds18. The use of fungal cultures has been considered as an environmental tool to remove organic pollutants such as polycyclic aromatic hydrocarbons, chlorinated and phenolic compounds, dyes, pharmaceutical compounds, among others19.
Lignin modifying enzymes
LMEs are oxidoreductases which catalyze the electron transfer from one substrate to another. LMEs act by generating free radicals that randomly attack the lignin molecule, breaking covalent bonds and releasing a range of phenolic compounds.
There are two main types of LMEs: peroxidases and laccases (phenol oxidases). The main LMEs are lignin peroxidase (LiP), manganese peroxidase (MnP), versatile peroxidase (VP) and laccases (Lac). In addition, these fungi secrete mediators of high molecular weight increasing the range of potentially biodegradable compounds. White-rot fungi start LMEs production during their secondary metabolism, since lignin oxidation provides no net energy to fungi 20. These enzymes are responsible for generating highly reactive and non-specific free radicals 21 that make them attractive for the development of advanced oxidation processes, where these enzymes may oxidize and degrade highly recalcitrant compounds. The main lignin-modifying enzymes are described below.
Laccase (Lac, E.C.1.10.3.2)
The enzyme laccase is a multi-copper oxidase that catalyzes one-electron oxidations by transferring one electron from four substrate molecules to one molecule of molecular oxygen which is reduced to water. Laccase shows low substrate specificity and can react with diphenols, aryl diamines, aminophenoles. The redox potential of laccases is in the range between 780-800 mV. However, in presence of a mediator, laccase is able to oxidize also non-phenolic molecules. It has been isolated from cultures of Aspergillus and thermophilic fungi such as Myceliophora thermophila, Chaemotium thermophilium, among others20.
Lignin peroxidase (LiP, E.C.1.11.1.14)
It was the first ligninolytic enzyme isolated in 1980’s decade from the fungus Phanerochaete chrysosporium22. It is a glycoprotein with molecular mass between 38 and 47 kDa with a distinctive feature of an unusually low pH optimum near 3. It is able to catalyze the oxidation of phenolic and aromatic compounds with a similar structure to lignin. LiP shows a classical peroxidase mechanism: it can react with phenolic aromatic substrates forming phenoxy radicals, but it is unique in its ability to oxidize substrates of high redox potential (up to 1.4 V)20.
Manganese peroxidase (MnP, E.C.1.11.1.13)
Manganese peroxidase is an extracellular enzyme which is considered the most widespread ligninolytic peroxidase produced by almost all white-rot basidiomycetes and by various litter-decomposing fungi20. MnP is a glycoprotein with molecular weights between 32 and 62.5 kDa. This enzyme has a similar catalytic cycle to other peroxidases involving a two-electron oxidation; however, MnP is able to oxidize Mn2+, resulting in the formation of diffusible oxidants (Mn3+) capable of penetrating the cell wall matrix and oxidizing mainly phenolic substrates23.
The catalytic cycle is initiated by binding of H2O2 to the native (ferric) enzyme and formation of an iron-H2O2 complex. Subsequent cleavage of the H2O2 oxygen-oxygen bond requires a two-electron transfer from the heme resulting in formation of MnP-I (Fe4+). Afterwards, the O-O bond is heterolytically cleaved and a H2O molecule released. Subsequent reduction proceeds through MnP-II (Fe4+). Mn2+ ion acts as one electron-donor for this porphyrin intermediate and is oxidized to Mn3+. The reduction of MnP-II proceeds in a similar way and another Mn3+ is formed from Mn2+, thereby leading to generation of native enzyme and release of the second water molecule. MnP is sensitive to high concentrations of H2O2 that cause reversible inactivation of the enzyme by forming MnP-III23, a catalytically inactive oxidation state but can be rescued by Mn3+ ions. This species can form complexes with organic acids, such as malonic or oxalic acid, secreted by the fungus in significant amounts that attack organic molecules non-specifically at location remote from the enzyme active site. These chelators could also accomplish other physiological functions; they enhance the dissociation of Mn2+ from the enzyme improving its activity, allow the fungus to control pH, sequester Ca2+ ions to increase the pore size of the plant cell wall and facilitate the penetration of the enzyme or react with O2 to form H2O2 useful for the enzyme activity23.
Versatile peroxidase (VP, E.C.1.11.1.16)
The enzyme VP is a peroxidase which combines the substrate specificity characteristics of the three other fungal peroxidases (MnP, LiP and Coprinopsis cinerea peroxidase). In this way, it is able to oxidize a variety of high and low redox potential substrates including Mn2+, phenolic and non-phenolic lignin dimers, α-keto-γ-thiomethyl-butyric acid (KTBA), veratryl alcohol- dimethoxybenzenes, different types of dyes (Reactive Black 5), substituted phenols and hydroquinones. VP is only produced by fungi from the genera Pleurotus, Bjerkandera and Lepista. The VP catalytic cycle includes two-electron oxidation of the resting peroxidase (VP, containing Fe3+) by hydrogen peroxide to yield compound I (C-IA, containing Fe4+-oxo and porphyrin cation radical), whose reduction in two one-electron reactions, producing Mn3+, results in the intermediate compound II (C-IIA, containing Fe4+-oxo after porphyrin reduction) and then the resting form of the enzyme. Compounds C-IB and C-IIB are involved in the oxidation of veratryl alcohol and other high redox potential aromatic compounds24.
Removal of EDCs by white rot fungi (WRF) and their lignin modifying enzymes (LMEs)
Although conventional biological treatment processes have been reported effective at reducing levels of some EDCs in wastewater and sewage, the low levels of these contaminants in wastewater effluents are still a major concern for the receiving environment and downstream users because EDCs exert physiological effects at very low concentrations. Therefore, post-treatments methods for the removal of these compounds are being investigated: (i) physical methods such as adsorption or membrane separation; (ii) chemical treatments, such as those based on oxidative catalysis, chlorination, ozonation and other advanced oxidation processes (AOPs). In general, these processes have high degradation rate but they have low selectivity and high costs. In addition, AOPs can be a cause of concern itself since they may render harmful by-products or transformation products which can have similar or increased estrogenicity of that of the parent compound.
An environmentally friendly alternative for the elimination of EDCs may be the use of microorganisms. Among the different possible microorganisms, white rot fungi appear to be a good choice since they have been reported to degrade a wide range of organic pollutants 19. Several authors have demonstrated the ability of different WRF not only to eliminate EDCs but also to reduce their estrogenic activity (Table 1).
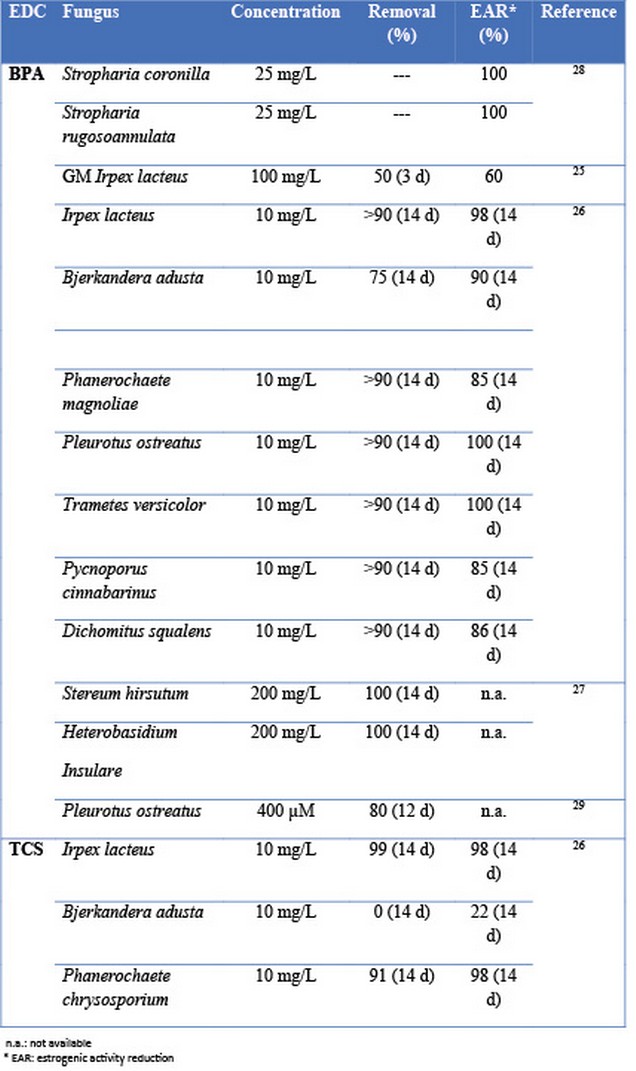
Table 1. Elimination of EDCs and reduction of estrogenicity by WRF
WRF were demonstrated to easily eliminate EDCs, reaching removal yields higher than 90%25,26 or even 100%26,27 when the initial concentration was within the order of mg/L. As a consequence, WRF were able to reduce the estrogenic activity up to values of 100%28. However, in the case of estrogenic compounds such as E1, E2 and EE2, the estrogenic activity reduction was lower (15-94%)26.
The main drawback of using white rot fungi for the degradation of organic pollutants is the necessity of working in aseptic conditions. Moreover, as shown in Table 1, the time required to eliminate 90% of the initial EDCs amount is between 10 and 14 days. As a consequence, the development of this alternative is costly and problematical.
The capability of this fungal class to remove pollutants usually is related to the production and secretion of lignin modifying enzymes. Several works have demonstrated that the use of LMEs is a good environmental tool for the elimination of organic pollutants EDCs (Table 2). Among the different enzymes, laccase is the most documented oxidative enzyme considered for the elimination of EDCs (Figure 3).
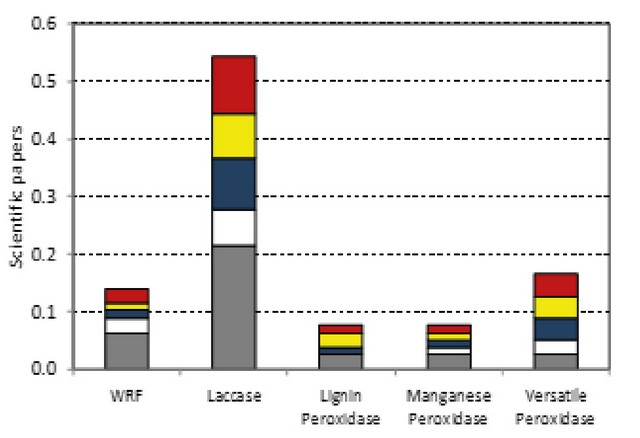
Figure 3. Impact of the in vivo and in vitro WRF processes in the elimination of EDCs.
BPA (grey), TCS (white), E1 (blue), E2 (yellow) and EE2 (red).
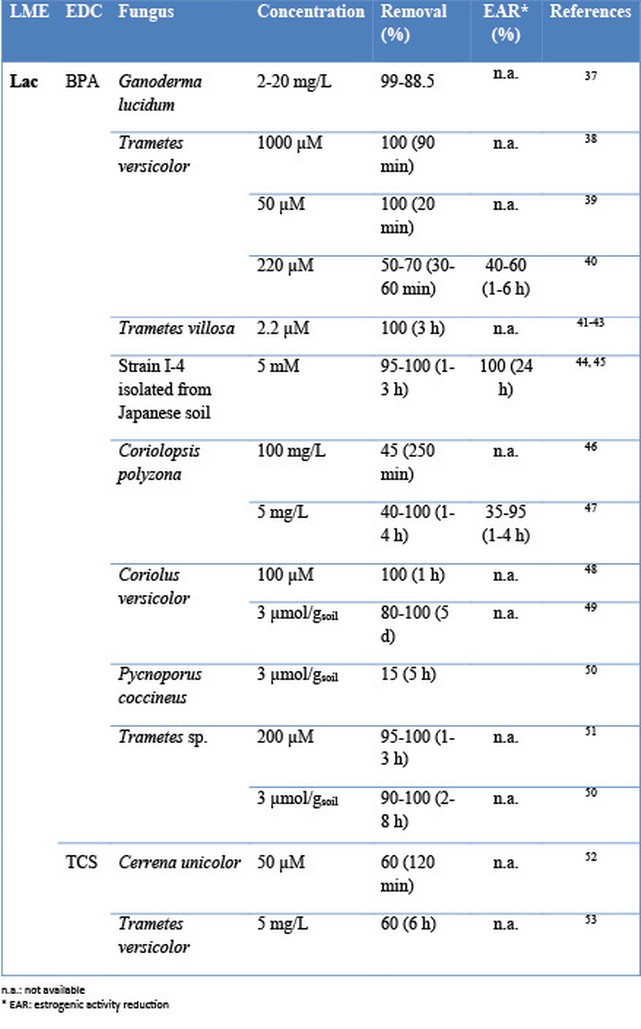
Table 2. Elimination of EDCs and reduction of estrogenicity by LMEs
The main advantage related to the use of this enzyme over other oxidases and peroxidases is its availability (e.g. DeniLite®, the commercial laccase preparation from Novozymes, Denmark) due to the scale-up of its production. However, one of its major disadvantages is that the redox potential of laccase is low (0.5 - 0.8 V) when compared with other ligninolytic enzymes such as peroxidases, 1.45-1.51 V 20 and it typically requires chemical mediators acting as the real oxidants, participating in the catalytic cycle of the enzyme33. Most of these mediators are environmentally unsafe (HBT, for example) and may have an important economical impact on the treatment. As a general approach, it can be established that removal yield values reached by using peroxidases are higher, between 90% and 100%34 than those observed when using laccases for the elimination of EDCs35.
From an economical point of view, an evaluation of the use of LMEs demonstrated that processes based on free peroxidase is economically competitive compared to photolysis, ozonolysis and Fenton processes36.
CONCLUSIONS
Existing traditional WWTPs are not able to eliminate EDCs completely, therefore the search of new technologies suitable for the treatment of wastewater containing them is a challenge to be addressed. The widespread technologies used for the elimination of EDCs are AOPs such as photocatalysis, photodegradation, sonolysis, ozonolysis, among others which have been demonstrated to get high elimination yields. However, these processes present low selectivity and high costs. Moreover, AOPs can be a cause of concern itself since they may render harmful by-products or transformation products which can have similar or increased estrogenicity of that of the parent compound.
A biological alternative may be the use of WRF and/or their LMEs. From an operational point of view, the use of LMEs in in vitro systems, compared to the use of WRF in in vivo systems, is easier, cheaper and faster. Among the LMEs, laccase is the most extensively studied enzyme for the degradation of BPA, TCS, E1, E2 and EE2. Nevertheless, peroxidases have higher redox potential and consequently they provide better results of EDC removal (in terms of yield and degradation rate).
Acknowledgements
R. Taboada-Puig would like to express his gratitude to the Yachay Tech University and to the Spanish Ministry of Science and Innovation for his financial support (BES-2008-006977) for his financial support. This work was financially supported by the Spanish Ministry of Economy and Competitiveness (CTQ2013-44762-R and CTQ2016-79461-R). The authors from the USC belong to the Galician Competitive Research Group GRC 2013-032 (program co-funded by FEDER) and to the strategic group CRETUS (AGRUP2015/02).
REFERENCES
1. USEPA, Special report on endocrine disruption. An effects assessment and analysis. In U.S. EPA: Washington, DC, 1997.
2. NTP-CERHR, Monograph on the potential human reproductive and developmental effects of bisphenol A. 2008.
3. C.G., Canada Gazette Part II. 2010, 144, (21).
4. Drewes, J. E.; Hemming, J.; Ladenburger, S. J.; Schauer, J.; Sonzogni, W., An assessment of endocrine disrupting activity changes during wastewater treatment through the use of bioassays and chemical measurements. Water Environment Research 2005, 77, (1), 12-23.
5. Alexander, H. C.; Dill, D. C.; Smith, L. W.; Guiney, P. D.; Dorn, P., Bisphenol A: acute aquatic toxicity. Environmental Toxicology and Chemistry 1988, 7, (1), 19-26.
6. Flint, S.; Markle, T.; Thompson, S.; Wallace, E., Bisphenol A exposure, effects, and policy: A wildlife perspective. Journal of Environmental Management 2012, 104, 19-34.
7. Jones, R. D.; Jampani, H. B.; Newman, J. L.; Lee, A. S., Triclosan: A review of effectiveness and safety in health care settings. American Journal of Infection Control 2000, 28, (2), 184-196.
8. C.G., Canada Gazette Part I. 2012, 146, (13).
9. Bedoux, G.; Roig, B.; Thomas, O.; Dupont, V.; Le Bot, B., Occurrence and toxicity of antimicrobial triclosan and by-products in the environment. Environmental Science and Pollution Research 2012, 19, (4), 1044-1065.
10. Yu, C.-P.; Chu, K.-H., Occurrence of pharmaceuticals and personal care products along the West Prong Little Pigeon River in east Tennessee, USA. Chemosphere 2009, 75, (10), 1281-1286.
11. Bester, K., Fate of triclosan and triclosan-methyl in sewage treatment plants and surface waters. Arch. Environ. Contam. Toxicol. 2005, 49, (1), 9-17.
12. Lopez-Avila, V.; Hites, R. A., Organic compounds in an industrial wastewater. Their transport into sediments. Environmental Science & Technology 1980, 14, (11), 1382-1390.
13. Heidler, J.; Halden, R. U., Mass balance assessment of triclosan removal during conventional sewage treatment. Chemosphere 2007, 66, (2), 362-369.
14. Johnson, A. C.; Belfroid, A.; Di Corcia, A., Estimating steroid oestrogen inputs into activated sludge treatment works and observations on their removal from the effluent. Science of the Total Environment 2000, 256, (2-3), 163-173.
15. Racz, L.; Goel, R. K., Fate and removal of estrogens in municipal wastewater. Journal of Environmental Monitoring 2010, 12, (1), 58-70.
16. Liu, Z.-h.; Kanjo, Y.; Mizutani, S., Removal mechanisms for endocrine disrupting compounds (EDCs) in wastewater treatment - physical means, biodegradation, and chemical advanced oxidation: A review. Science of the Total Environment 2009, 407, (2), 731-748.
17. Schwarze, F.; Baum, S.; Fink, S., Dual modes of degradation by Fistulina hepatica in xylem cell walls of Quercus robur. Mycological Research 2000, 104, 846-852.
18. Mester, T.; Tien, M., Oxidation mechanism of ligninolytic enzymes involved in the degradation of environmental pollutants. International Biodeterioration & Biodegradation 2000, 46, (1), 51-59.
19. Kulikova, N. A.; Klein, O. I.; Stepanova, E. V.; Koroleva, O. V., Use of Basidiomycetes in Industrial Waste Processing and Utilization Technologies: Fundamental and Applied Aspects (Review). Applied Biochemistry and Microbiology 2011, 47, (6), 565-579.
20. Wesenberg, D.; Kyriakides, I.; Agathos, S. N., White-rot fungi and their enzymes for the treatment of industrial dye effluents. Biotechnology Advances 2003, 22, (1-2), 161-187.
21. Kersten, P.; Cullen, D., Extracellular oxidative systems of the lignin-degrading Basidiomycete Phanerochaete chrysosporium. Fungal Genetics and Biology 2007, 44, (2), 77-87.
22. Tien, M.; Kirk, T. K., Lignin peroxidase of Phanerochaete chrysosporium. Method Enzymol 1988, 161, 238-249.
23. Wong, D. W. S., Structure and Action Mechanism of Ligninolytic Enzymes. Applied Biochemistry and Biotechnology 2009, 157, (2), 174-209.
24. Perez-Boada, M.; Ruiz-Duenas, F. J.; Pogni, R.; Basosi, R.; Choinowski, T.; Martinez, M. J.; Piontek, K.; Martinez, A. T., Versatile peroxidase oxidation of high redox potential aromatic compounds: Site-directed mutagenesis, spectroscopic and crystallographic investigation of three long-range electron transfer pathways. Journal of Molecular Biology 2005, 354, (2), 385-402.
25. Kum, H.; Kim, M. K.; Choi, H. T., Degradation of endocrine disrupting chemicals by genetic transformants in Irpex lacteus with an inducible laccase gene of Phlebia tremellosa. Biodegradation 2009, 20, (5), 673-678.
26. Cajthaml, T.; Kresinova, Z.; Svobodova, K.; Moder, M., Biodegradation of endocrine-disrupting compounds and suppression of estrogenic activity by ligninolytic fungi. Chemosphere 2009, 75, (6), 745-750.
27. Lee, S. M.; Koo, B. W.; Lee, S. S.; Kim, M. K.; Choi, D. H.; Hong, E. J.; Jeung, E. B.; Choi, I. G., Biodegradation of dibutylphthalate by white rot fungi and evaluation on its estrogenic activity. Enzyme and Microbial Technology 2004, 35, (5), 417-423.
28. Kabiersch, G.; Rajasarkka, J.; Ullrich, R.; Tuomela, M.; Hofrichter, M.; Virta, M.; Hatakka, A.; Steffen, K., Fate of bisphenol A during treatment with the litter-decomposing fungi Stropharia rugosoannulata and Stropharia coronilla. Chemosphere 2011, 83, (3), 226-232.
29. Hirano, T.; Honda, Y.; Watanabe, T.; Kuwahara, M., Degradation of bisphenol a by the lignin-degrading enzyme, manganese peroxidase, produced by the white-rot basidiomycete, Pleurotus ostreatus. Bioscience Biotechnology and Biochemistry 2000, 64, (9), 1958-1962.
30. Hundt, K.; Martin, D.; Hammer, E.; Jonas, U.; Kindermann, M. K.; Schauer, F., Transformation of triclosan by Trametes versicolor and Pycnoporus cinnabarinus. Applied and Environmental Microbiology 2000, 66, (9), 4157-4160.
31. Tamagawa, Y.; Hirai, H.; Kawai, S.; Nishida, T., Removal of estrogenic activity of endocrine-disrupting genistein by ligninolytic enzymes from white rot fungi. Fems Microbiology Letters 2005, 244, 93-98.
32. Blanquez, P.; Guieysse, B., Biodegradation of estrogenic compounds by white rot fungi. J. Biotechnol. 2007, 131, (2), S244-S245.
33. Lloret, L.; Eibes, G.; Lu-Chau, T. A.; Moreira, M. T.; Feijoo, G.; Lema, J. M., Laccase-catalyzed degradation of anti-inflammatories and estrogens. Biochemical Engineering Journal 2010, 51, (3), 124-131.
34. Taboada-Puig, R.; Eibes, G.; Lloret, L.; Lu-Chau, T. A.; Feijoo, G.; Moreira, M. T.; Lema, J. M., Fostering the action of versatile peroxidase as a highly efficient biocatalyst for the removal of endocrine disrupting compounds. New Biotechnology 2016, 33, (1), 187-195.
35. Lloret, L.; Eibes, G.; Feijoo, G.; Moreira, M. T.; Lema, J. M., Application of response surface methodology to study the removal of estrogens in a laccase-mediated continuous membrane reactor. Biocatalysis and Biotransformation 2013, 31, (4), 197-207.
36. Lopez, C.; Teresa Moreira, M.; Feijoo, G.; Manuel Lema, J., Economic comparison of enzymatic reactors and advanced oxidation processes applied to the degradation of phenol as a model compound. Biocatalysis and Biotransformation 2011, 29, (6), 344-353.
37. Liao, C.-S.; Yuan, S.-Y.; Hung, B.-H.; Chang, B.-V., Removal of organic toxic chemicals using the spent mushroom compost of Ganoderma lucidum. Journal of Environmental Monitoring 2012, 14, (7), 1983-1988.
38. Nicolucci, C.; Rossi, S.; Menale, C.; Godjevargova, T.; Ivanov, Y.; Bianco, M.; Mita, L.; Bencivenga, U.; Mita, D. G.; Diano, N., Biodegradation of bisphenols with immobilized laccase or tyrosinase on polyacrylonitrile beads. Biodegradation 2011, 22, (3), 673-683.
39. Georgieva, S.; Godjevargova, T.; Mita, D. G.; Diano, N.; Menale, C.; Nicolucci, C.; Carratelli, C. R.; Mita, L.; Golovinsky, E., Non-isothermal bioremediation of waters polluted by phenol and some of its derivatives by laccase covalently immobilized on polypropylene membranes. Journal of Molecular Catalysis B-Enzymatic 2010, 66, (1-2), 210-218.
40. Tsutsumi, Y.; Haneda, T.; Nishida, T., Removal of estrogenic activities of bisphenol A and nonylphenol by oxidative enzymes from lignin-degrading basidiomycetes. Chemosphere 2001, 42, (3), 271-276.
41. Fukuda, T.; Uchida, H.; Suzuki, M.; Miyamoto, H.; Morinaga, H.; Nawata, H.; Uwajima, T., Transformation products of bisphenol A by a recombinant Trametes vilosa laccase and their estrogenic activity. Journal of Chemical Technology and Biotechnology 2004, 79, (11), 1212-1218.
42. Fukuda, T.; Uchida, H.; Takashima, Y.; Uwajima, T.; Kawabata, T.; Suzuki, M., Degradation of bisphenol a by purified laccase from Trametes villosa. Biochemical and Biophysical Research Communications 2001, 284, (3), 704-706.
43. Uchida, H.; Fukuda, T.; Miyamoto, H.; Kawabata, T.; Suzuki, M.; Uwajima, T., Polymerization of Bisphenol A by Purified Laccase from Trametes villosa. Biochemical and Biophysical Research Communications 2001, 287, (2), 355-358.
44. Saito, T.; Hong, P.; Kato, K.; Okazaki, M.; Inagaki, H.; Maeda, S.; Yokogawa, Y., Purification and characterization of an extracellular laccase of a fungus (family Chaetomiaceae) isolated from soil. Enzyme and Microbial Technology 2003, 33, (4), 520-526.
45. Saito, T.; Kato, K.; Yokogawa, Y.; Nishida, M.; Yamashita, N., Detoxification of bisphenol A and nonylphenol by purified extracellular laccase from a fungus isolated from soil. Journal of Bioscience and Bioengineering 2004, 98, (1), 64-66.
46. Cabana, H.; Alexandre, C.; Agathos, S. N.; Jones, J. P., Immobilization of laccase from the white rot fungus Coriolopsis polyzona and use of the immobilized biocatalyst for the continuous elimination of endocrine disrupting chemicals. Bioresource Technology 2009, 100, (14), 3447-3458.
47. Cabana, H.; Jiwan, J. L. H.; Rozenberg, R.; Elisashvili, V.; Penninckx, M.; Agathos, S. N.; Jones, J. P., Elimination of endocrine disrupting chemicals nonylphenol and bisphenol A and personal care product ingredient triclosan using enzyme preparation from the white rot fungus Coriolopsis polyzona. Chemosphere 2007, 67, (4), 770-778.
48. Okazaki, S.-y.; Michizoe, J.; Goto, M.; Furusaki, S.; Wariishi, H.; Tanaka, H., Oxidation of bisphenol A catalyzed by laccase hosted in reversed micelles in organic media. Enzyme and Microbial Technology 2002, 31, (3), 227-232.
49. Tanaka, T.; Yamada, K.; Tonosaki, T.; Konishi, T.; Goto, H.; Taniguchi, M., Enzymatic degradation of alkylphenols, bisphenol A, synthetic estrogen and phthalic ester. Water Sci Technol 2000, 42, (7-8), 89-95.
50. Tanaka, T.; Tonosaki, T.; Nose, M.; Tomidokoro, N.; Kadomura, N.; Fujii, T.; Taniguchi, M., Treatment of model soils contaminated with phenolic endocrine-disrupting chemicals with laccase from Trametes sp in a rotating reactor. J Biosci Bioeng 2001, 92, (4), 312-316.
51. Michizoe, Y.; Ichinose, H.; Kamiya, N.; Maruyama, T.; Goto, M., Biodegradation of phenolic environmental pollutants by a surfactant-laccase complex in organic media. Journal of Bioscience and Bioengineering 2005, 99, (6), 642-647.
52. Songulashvili, G.; Jimenez-Tobon, G. A.; Jaspers, C.; Penninckx, M. J., Immobilized laccase of Cerrena unicolor for elimination of endocrine disruptor micropollutants. Fungal biology 2012, 116, (8), 883-9.
53. Cabana, H.; Ahamed, A.; Leduc, R., Conjugation of laccase from the white rot fungus Trametes versicolor to chitosan and its utilization for the elimination of triclosan. Bioresource Technology 2011, 102, (2), 1656-1662.
54. Inoue, Y.; Hata, T.; Kawai, S.; Okamura, H.; Nishida, T., Elimination and detoxification of triclosan by manganese peroxidase from white rot fungus. Journal of Hazardous Materials 2010, 180, (1-3), 764-767.
55. Murugesan, K.; Chang, Y.-Y.; Kim, Y.-M.; Jeon, J.-R.; Kim, E.-J.; Chang, Y.-S., Enhanced transformation of triclosan by laccase in the presence of redox mediators. Water Research 2010, 44, (1), 298-308.
56. Tanaka, T.; Tamura, T.; Ishizaki, Y.; Kawasaki, A.; Kawase, T.; Teraguchi, M.; Taniguchi, M., Enzymatic treatment of estrogens and estrogen glucuronide. Journal of Environmental Sciences-China 2009, 21, (6), 731-735.
57. Auriol, M.; Filali-Meknassi, Y.; Tyagi, R. D.; Adams, C. D., Laccase-catalyzed conversion of natural and synthetic hormones from a municipal wastewater. Water Research 2007, 41, 3281-3288.
58. Auriol, M.; Filali-Meknassi, Y.; Adams, C. D.; Tyagi, R. D.; Noguerol, T. N.; Pina, B., Removal of estrogenic activity of natural and synthetic hormones from a municipal wastewater: Efficiency of horseradish peroxidase and laccase from Trametes versicolor. Chemosphere 2008, 70, (3), 445-452.
59. Wang, J.; Majima, N.; Hirai, H.; Kawagishi, H., Effective Removal of Endocrine-Disrupting Compounds by Lignin Peroxidase from the White-Rot Fungus Phanerochaete sordida YK-624. Current Microbiology 2012, 64, (3), 300-303.
60. Kimura, M.; Michizoe, J.; Oakazaki, S.; Furusaki, S.; Goto, M.; Tanaka, H.; Wariishi, H., Activation of lignin peroxidase in organic media by reversed micelles. Biotechnology and Bioengineering 2004, 88, (4), 495-501.
61. Mao, L.; Huang, Q.; Lu, J.; Gao, S., Ligninase-Mediated Removal of Natural and Synthetic Estrogens from Water: I. Reaction Behaviors. Environmental Science & Technology 2009, 43, (2), 374-379.
62. Tamagawa, Y.; Yamaki, R.; Hirai, H.; Kawai, S.; Nishida, T., Removal of estrogenic activity of natural steroidal hormone estrone by ligninolytic enzymes from white rot fungi. Chemosphere 2006, 65, (1), 97-101.
63. Suzuki, K.; Hirai, H.; Murata, H.; Nishida, T., Removal of estrogenic activities of 17 beta-estradiol and ethinylestradiol by ligninolytic enzymes from white rot fungi. Water Res 2003, 37, (8), 1972-1975.
64. Eibes, G.; Debernardi, G.; Feijoo Costa, G.; Moreira Vilar, M. T.; Lema Rodicio, J. M., Oxidation of pharmaceutically active compounds by a ligninolytic fungal peroxidase. Biodegradation 2010.
Recibido: Enero 2017
Aprobado: febrero 2017
Roberto Taboada-Puig1*, Maria Teresa Moreira2, Spiros N. Agathos1, Juan M. Lema2
1School of Biological Sciences and Engineering, Yachay Tech University, Hacienda San José s/n y Proyecto Yachay, San Miguel de Urcuquí – Ecuador
2Department of Chemical Engineering, Institute of Technology, University of Santiago de Compostela, E-15782, Santiago de Compostela, Spain.
Maria Teresa Moreira ([email protected]), Spiros N. Agathos ([email protected]), Juan M. Lema ([email protected])
*Corresponding author:
Roberto Taboada Puig, PhD
1School of Biological Sciences and Engineering, Yachay Tech University, Hacienda San José s/n y Proyecto Yachay, San Miguel de Urcuquí – Ecuador