2023.08.03.22
Files > Volume 8 > Vol 8 No 3 2023
Nutritional supplement of FOS enhances growth and immune system in tilapia larvae (Oreochromis niloticus)

Antonio Morales1 , Daniela F. Moro1, Adrian Rodriguez1, Liz Hernández1, Amanda Comellas1, Fidel Herrera1, Osmany Gonzalez1, Enrique Pérez Cruz2, Mario Pablo Estrada1, Rebeca Martinez1*
, Daniela F. Moro1, Adrian Rodriguez1, Liz Hernández1, Amanda Comellas1, Fidel Herrera1, Osmany Gonzalez1, Enrique Pérez Cruz2, Mario Pablo Estrada1, Rebeca Martinez1*
1 Metabolic Modifiers for Aquaculture, Agricultural Biotechnology Department, Center for Genetic Engineering and Biotechnology, Havana 10600, Cuba.
2 Diagnostic Division, Center for Genetic Engineering and Biotechnology, Sancti Spiritus, Cuba.
* Correspondence author: [email protected]
Available from: http://dx.doi.org/10.21931/RB/2023.08.03.22
ABSTRACT
Aquaculture is an essential economic activity worldwide, supplying the requirements of various nations. The larval stage is one of the most critical periods during the production cycle in aquaculture. To contribute to amplifying the knowledge related to viable methods to enhance this activity, the present study investigated the effects of prebiotic fructooligosaccharide (FOS) and the synthetic peptide GHRP-6 in the growth of fish, as well as its repercussions on a series of digestive enzyme activities and immune response parameters. Tilapias (Oreochromis niloticus) larvae were divided into three groups: group 1, fed with pre-formulated feed; group 2, fed with feed + FOS; group 3, fed with feed + GHRP-6. We measured the gain in size and weight at 72 and 87 days after starting the diets. At the end of treatment, we continue feeding the animals with a standard diet using the pre-formulated feed. We measured immune system parameters and digestive enzyme activities on larvae homogenate, focusing on lysozyme and antiprotease activity and α-amylase, trypsin and esterase activity, respectively. Results show a significant increment in size and weight in those fish fed with feed + FOS. Meanwhile, the primary enzyme activity was detected in those fish fed with feed + GHRP-6. On rough terms, both food combinations stimulated the immune system.
Our study suggests that FOS and GHRP-6 could be desirable additives to incorporate into the tilapia larvae feeding and increase production efficiency.
Keywords: aquaculture, digestive enzymes activity, FOS, GHRP-6, immune system, larvae
INTRODUCTION
Current times are moved by the need to create a sustainable future, in which adjusting to the 2030 Agenda for Sustainable Development has become challenging. In this scenario, aquaculture is the fastest-growing food production sector all over the world1. Consequently, aquaculture should develop actions as responses to Sustainable Development Goals (SDGs).
When involved in feeding animals, protein and oil from marine sources have been the basis of aquaculture diets, which nowadays become unsustainable2. Therefore, optimizing the main complements of diet is necessary to enhance fish's growth and healthcare. Due to its low cost, fast growth, high protein level and other peculiar specifications, Nile tilapia (Oreochromis niloticus) is an essential species in this economic sector1,3.
One of the strategies to improve the effectiveness of diets is the use of dietary supplements, such as prebiotics2, such as mannan- oligosaccharide (MOS), fructo-oligosaccharide (FOS) and galacto-oligosaccharide (GOS). Prebiotics are generally specially fermented by anaerobic bacteria in the distal intestine of the host, resulting in various secondary metabolites, mainly short-chain fatty acids4,5. Prebiotics stimulate the activity of commensal bacteria that are beneficial to the host, increasing its growth and microbial activity of the digestive tract, enhancing the resistance to stress and pathogens, and thus, the immune system6.
Consequently, prebiotics have been used in aquatic animal diets for decades. However, the efficacy of a prebiotic in aquatic animals may vary depending on the prebiotic itself, the host species, its feeding habits, the basal diet type and the intestinal microbiome7,8. Also, prebiotics have shown to be a promise as eco-friendly additives to improve growth performance, feed utilization, immunological status and disease resistance in aquaculture9.
FOS is known to be composed of short and medium chains of beta-linked fructose units ending with glucose10. Previous studies have shown that 0.4% dietary FOS significantly improved the growth performance, intestinal short-chain fatty acids production and hepatopancreatic condition in the freshwater prawn (Macrobrachium rosenbergii)10. In blunt snout bream11 and juvenile ovate pompano (Trachinotus ovatus)12, 4 g/kg, FOS inclusion was reported with the highest levels of growth parameters. In general, the potentials of dietary FOS in aquaculture have been mentioned to promote growth performance, digestive enzyme activity, immune system, antioxidant capacity and beneficial gut microbiota13.
Growth Hormone-Releasing Peptide 6 (GHRP-6) is a six amino acid synthetic peptide commonly used in our laboratory on different growth and antimicrobial resistance studies in aquatic organisms14,15,16. Its effects on immune system parameters of teleost fish have been previously addressed in our research group. Lectin titers and intraepithelial lymphocytes increased when the peptide was orally administered to tilapia larvae14. Also, co-administering this peptide with different antigens via the intraperitoneal route stimulated antibody production in juvenile tilapia15. GHRP-6 stimulated the antiprotease and lysozyme levels in tilapia larvae homogenates; the stimulation of these activities has been related to a higher degree of disease resistance in fish17,18,19.
Previous studies in aquaculture rarely explore the beneficial effects of 0.4% dietary FOS supplementation on growth and related parameters centered in tilapia. On the other hand, studies about GHRP-6 effects, specifically on the digestive enzyme in tilapia larvae, have not been found in the literature either. Thus, the present study aimed to evaluate the efficiency of FOS and GHRP-6 diets on growth, immune system and digestive enzyme activities on tilapia larvae homogenate.
MATERIALS AND METHODS
Ethical statement
The Ethics Committee previously approved all fish manipulation for the care and use of scientific animals of the Center for Genetic Engineering and Biotechnology, Havana, Cuba. The experimental protocols and methodologies followed the EU Directive 84 2010/63/EU for animal experiments.
Experimental design and diets
Fructo-oligosaccharide (FOS: 95% purity) (Pérez, E. R. et al., 2021) was provided by Sancti Spíritus Genetic Engineering and Biotechnology Center, Cuba. GHRP-6 (His-(D-Trp)-Ala-Trp-(D-Phe)-Lys-NH2, MW = 872.44 Da) with a purity > 99% was provided by Sigma-Aldrich, USA. Three experimental diets were prepared: pre-formulated feed (basal diet), pre-formulated feed + FOS at 0.4%, and pre-formulated feed + GHRP-6 (500ug/Kg)14. The pre-formulated feed used in this study is a commercial dry diet for fish from CENPALAB, Havana, Cuba. All experimental diets were preserved at 4°C until its use. The dietary formulation of FOS on the experimental diets is presented in Table 1.
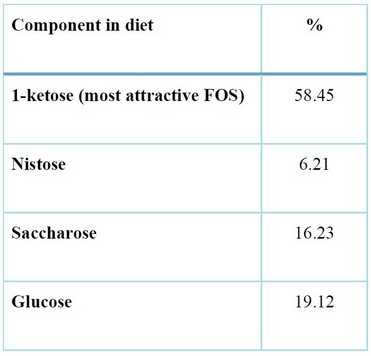
Table 1. FOS formulation on the experimental diets
Fish administration
Tilapia larvae were purchased from the Aquaculture Research Station at the Center for the Genetic Engineering and Biotechnology (CIGB), Havana, Cuba. Larvae were kept alive in aerated freshwater in 120 L tanks under a 12 h light: 12 h dark photoperiod. Water temperature was maintained at 26 - 28 °C. Before the feeding trial, all fish were acclimatized to experimental diets and conditions for a week. Five hundred animals per experimental group (250 larvae per group, twice replicated) were randomly separated into the three experimental groups physically delimited by each corresponding tank. Fish were fed thrice daily (8.00h, 12.00h and 16.00h). The feeding trial lasted for 90 days. The Center for Genetic Engineering and Biotechnology Ethics Committee, Havana, Cuba, previously approved all animal experiments. The described work followed the EU Directive 2010/63/EU for animal experiments.
RESULTS
Growth parameters and sample collection
Size and growth of tilapia larvae were measured at 72 and 87 days from the start of the feeding process. The weight gain (WG) was calculated following the below formula:
WG (%) =100× (W2 −W1) ∕ W1
Where W1 and W2 are the initial and final weights of fish.
Samples were taken 90 days after the start of the feeding process to study the immune system modulation. At the same time, intestine samples were collected to measure the digestive enzyme activities. Every model was stored at −70 °C until its use.
Digestive enzyme activities
Determination of α-amylase activity
The substrate used to determine α-amylase activity was 2-chloro-4-nitrophenyl-α-maltotrioside (CNP-G3)20. The activity was measured using a buffer in our laboratory (50mM MES; 1mM CaCl; 70mM NaCl; 2.25mM CNP-G3; pH 5.5). The assay was performed in 96-well microplates. The reaction mixture comprised 10 μL of extract (1/100) and 200μL of buffer with CNP-G3. The mixture was incubated for 60 seconds at 37 °C20. Released para nitrophenol (CNP) was detected kinetically every 15 seconds for 4 minutes at 405 nm and 37 °C using a Biotek ELx808IU microplate reader. Initial velocity was determined as the slope of the linear portion of the absorbance vs. time graph using KC4 software (BioTek). Amylase activity was expressed in units per milliliter (U/mL). The enzyme activity unit (U) was taken as necessary to increase one absorbance unit per minute.
Determination of trypsin-like activity
The trypsin-like activity of the enzymes was determined using 1.25 mM BApNA in 0.2 M Tris-HCl, 20 mM CaCl2, and pH 8.4. BApNA stock solution (125 mM) was prepared in DMSO and brought to working concentration through dilution with buffer before analysis. The assay was performed in 96-well microplates, and the reaction mixture consisted of 10 µL of extract and 200 µL of substrate. Enzyme activity was measured continuously by p-nitroaniline release at 405 nm in an ELx808 IU reader, BioTek. Analyzes were performed in triplicate; data were recorded using BioTek's KC4 software, and the enzymatic activity was calculated as follows:
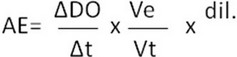
Where AE: enzymatic activity, ΔDO/Δt: absorbance variation over time, Ve: volume of enzyme extract in the assay, Vt: total volume in the assay, dil.: extract dilution. Enzyme activity was expressed as U/mL.
Determination of esterase activity
Esterase activity was measured by hydrolysis of 0.3 mM p-nitrophenyl butyrate (p-NPB) according to Gilham and Lehner (2005)21, with slight modifications. Stock solutions for these substrates (100 mM) were prepared in CH2Cl2 and diluted with 20 mM Tris-HCl, 150 mM NaCl, and pH 8.0 before assays. These consisted of 20 µL of extract and 200 µL of the substrate in buffer. The activity was measured in a 96-well microplate, kinetically following the release of p-nitrophenol at 405 nm in a Multiscan EX reader (Thermolab Systems).
Immune parameters
Antiprotease activity
Antiprotease activity (trypsin activity % of inhibition) was performed as Hernández L et al. (2021)19 described. Briefly, 20 µL of the homogenates were incubated with an identical volume of standard trypsin solution (Sigma-Aldrich, USA) (5 mg/mL) for 10 min at 22 °C. Subsequently, 200 µL of 0.1 M phosphate buffer (pH 7.0) and 250 µL 2% azocasein (Sigma-Aldrich, USA) were added and incubated for 1 h at 22 °C. 500 µL of 10% trichloroacetic acid was added for its incubation during 30 min at 22 °C. The mixture was centrifuged at 6000 g for 5 min, and 100 µL of the resulting supernatant was transferred to a 96-well microplate containing 100 µL/well of 1 M NaOH and read at 450 nm. Buffer replaced the homogenates for positive control, while for the negative control, buffer replaced both homogenates and trypsin. The percentage of inhibition of trypsin activity by each sample was calculated by comparing it to the 100% control sample. All the samples collected were analyzed in triplicates.
Lysozyme activity
The lysozyme activity in larvae homogenates samples was measured using a method described by Hernández L et al. (2021)19. Briefly, in a 96-well microarray, 100 μL of samples in four two-fold serial dilutions in phosphate buffer (0.05 M, pH 6.2) was mixed with 100 μL of a 0.4 mg/mL suspension of M. lysodeikticus (Sigma-Aldrich, USA) in phosphate buffer. Microtray was incubated at 22°C and read at 450nm at 0, 15, 30, and 60 min. For the positive control, homogenates were replaced by serial dilutions starting at 10 μg/mL of hen egg-white lysozyme; for the negative control, buffer replaced homogenates. A unit of lysozyme activity was defined as the amount of sample causing a decrease in the OD reading of 0.001 min. Lysozyme concentration was expressed as U per mL of sample (U mL-1).
Growth parameters
Growth-promoting effects were evaluated by measuring the body weight and size increase of all individuals in the population for 72 and 87 days.
The effect of FOS and GHRP-6 on larval growth was evaluated by body weight and size increase. As shown in Figure 1, the FOS and GHRP-6 supplementation at 4 g/kg significantly increased both weight and size of tilapia larvae (p < 0.05). In both cases, for 87 days, fish showed a higher gain in the evaluated parameters concerning the measurement made at 75 days. It is also evident that a higher modulation on these parameters by the effect of FOS in diet, compared with the modulation shown by GHRP-6 intake, under the specific conditions established in this experiment.
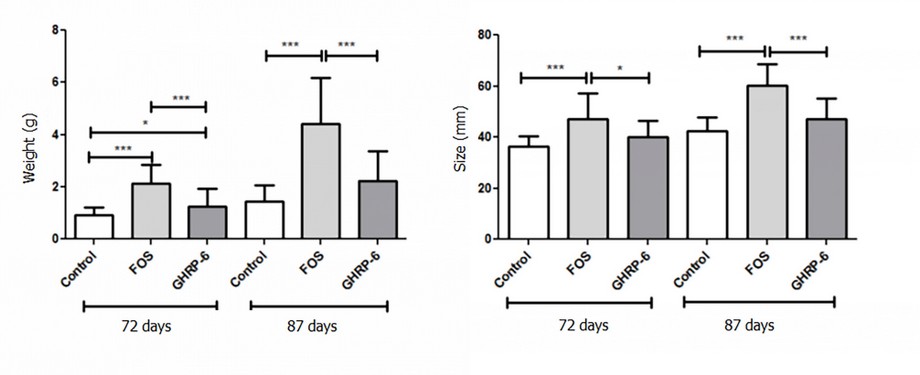
Figure 1. Growth experiment in tilapia larvae fed with GHRP-6 and FOS. Three experimental diets were administrated to fish: pre-formulated feed (control group), pre-formulated feed + FOS at 0.4%, and pre-formulated feed + GHRP-6 (500ug/Kg). GHRP-6 and FOS growth-promoting effects were evaluated by measuring the body weight (A) and size (B) at 72 and 87 days from the start of the feeding process. Data are represented as mean + standard deviation. The Kruskal analyzed data–Wallis test followed by Dunn's Multiple Comparison Test. Asterisks denote statistically significant differences: (*) p < 0.05; (**) p < 0.01; (***) p < 0.001.
Intestinal enzyme activities in larvae homogenates
The α-amylase, trypsin and esterase activity were measured on larvae homogenates. As shown in Figure 2, GHRP-6 treatment showed an increase in the α-amylase, trypsin and esterase activity concerning the control group (p < 0.05). On the other hand, statistically significant decreases were detected in FOS supplementation for the α-amylase and trypsin activity concerning the control group (p > 0.05). The esterase activity treated with FOS did not show any difference when compared with the control group.
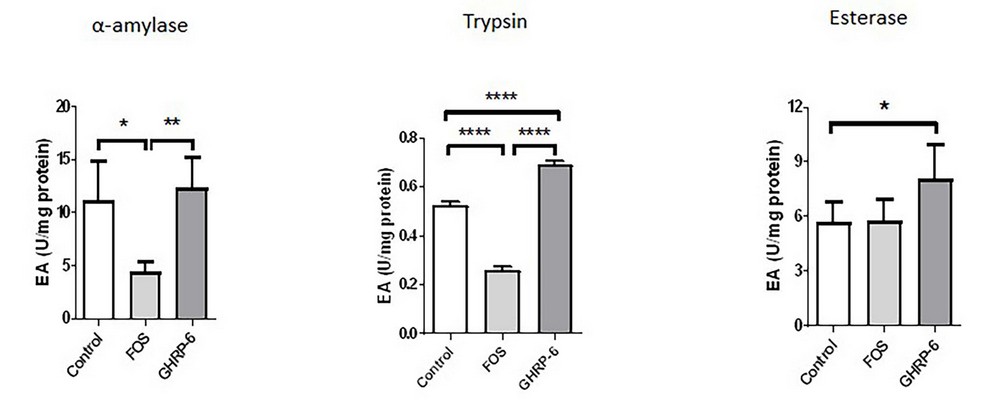
Fig. 2. Digestive enzyme activities in tilapia larvae at 90 days from the beginning of the experiment: α-amylase (A), trypsin (B), and esterase (C). Three experimental diets were suministrated to fish: pre-formulated feed (control group), pre-formulated feed + FOS at 0.4%, and pre-formulated feed + GHRP-6 (500ug/Kg). Data are represented as mean + standard deviation. Asterisks represent statistically significant differences: (*) p < 0.05; (**) p < 0.01; (***) p < 0.001 according to a T-student test.
Immune parameters in larvae homogenates
The trypsin inhibition and lysozyme activity were evaluated to obtain an approach to the effects of GHRP-6 and FOS supplementation on immune parameters in tilapia larvae. As shown in Figure 3, FOS supplementation significantly increased both antiprotease activity and lysozyme activity in larvae homogenates (p < .05). Meanwhile, on GHRP-6 diets, fish only showed significant differences in the lysozyme activity (p < .05), while the antiprotease activity, no significant differences were detected (p > .05), under the specific conditions established in this experiment.
Lysozyme activity
Antiprotease activity
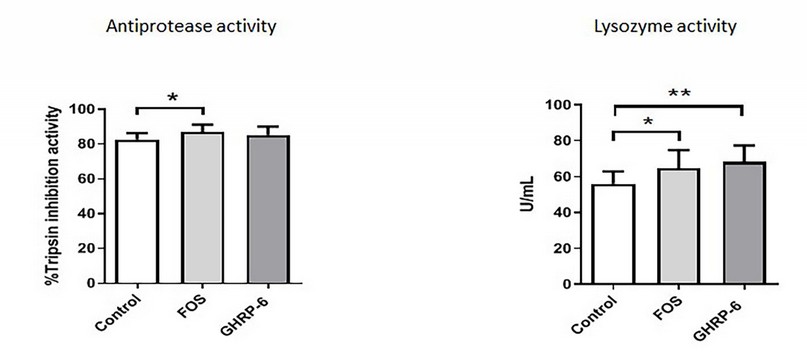
Figure. 3. Immune parameters in tilapia larvae at 90 days from the beginning of the experiment: trypsin activity % of inhibition (A) and lysozyme activity (B). Three experimental diets were suministrated to fish: pre-formulated feed (control group), pre-formulated feed + FOS at 0.4%, and pre-formulated feed + GHRP-6 (500ug/Kg). Asterisks represent statistically significant differences: (*) p < 0.05; (**) p < 0.01; (***) p < 0.001 according to a T-student test.
DISCUSSION
In this study, dietary supplementation of GHRP-6 and FOS on two independent diets significantly promoted the weight and size of tilapia (Oreochromis niloticus), indicating the positive effect of both molecules in the somatic growth performance.
According to the literature, GHSs such as GHRP-6 are valuable as growth enhancement molecules. They effectively stimulate the release of endogenous growth hormone as a physiological response, with no side effects or toxicity22. Else, it has been demonstrated that intraperitoneal administration of GHRP-6 to juvenile tilapia increased serum GH and transcription of IGF-I in the liver, with the GH-IGF system being the key growth promoter in vertebrates14. Thus, GHRP-6 increased body weight when administered by immersion baths to tilapia larvae, which enhanced their non-specific immunity14,22.
On the other hand, prebiotics such as FOS are functional dietary supplements. Previous studies have shown prebiotics to be a promise on the immune response stimulation, as well as on the resistance to pathogens and the improvement of growth performance and feed utilization23. Due to a lack of β-glucosidases, the digestive system of mammals and fishes cannot hydrolyze the β-(2-1) glycosidic linkages on FOS structure24. However, these animals have colonic bacteria expressing this enzyme, through which FOS can be fermented25. FOS treatment has been shown to potentiate specific intestinal microbiota, improving protein digestion and intestinal morphology24,26. In addition, it contributes to improving growth, feed efficiency and disease resistance in the animals. Thus, it is clarified that both GHRP-6 and FOS diets enhance the somatic growth of fish.
The present study also evaluates the activity of three digestive enzymes under the influence of both GHRP-6 and FOS diets on tilapia larvae homogenates. We measured the α-amylase, trypsin and esterase activity. On the GHRP-6 treatment, each enzyme activity showed a significant increase concerning the control group. GHRP-6 has proved to enhance feeding efficiency27.
It has been demonstrated that GHRP-6 increases feed intake in larval and juvenile teleosts14,27. This could lead to comprehending this peptide as a metabolism and growth promoter. At the time, this could be related to the potentiation of the enzymatic activity of α-amylase, trypsin and esterase. It is known that, meanwhile, α-amylase catalyzes the hydrolysis of α-glycosidic bonds on high-weight polysaccharides, and esterase catalyzes the hydrolysis of alcoholic and acid compounds, trypsin is fundamental to food protein absorption; all of them decisive processes on the food intake in the organisms. Other studies analyzing the effects of ghrelin analogs and weather on tilapia or other species, such as golden carp (Carassius auratus), had demonstrated the potentiation of feed intake using different procedures.
On the FOS supplementation, significant decreases were detected for the α-amylase and trypsin activity concerning the control group, while the esterase activity showed a non-significant increase propensity.
About these findings, other studies on different fish species also report a non-significant increase in effects on feed utilization by FOS diets6,31,32. Intestinal enzyme studies on a fish-fed supplemented diet with FOS showed no significant effects compared to control 11,12,33. In contrast, a study on the amylase and lipase recorded significant increases in its enzymatic activities when feeding Caspian roach (Rutilus rutilus) with 1, 2 and 3% FOS inclusion diets6,23. According to Wu et al. 201311, these differences may be related to fish species, FOS inclusion level, FOS resource, fish size, culture condition, feeding duration, and FOS composition.
However, the fact that digestive enzyme activity is positively correlated with intestinal digestion and absorption facilitates the growth performance in fish34, which has been previously demonstrated in this study. In addition, the improvement of somatic growth by FOS diets could be attributed to dietary FOS indirectly modifying fish metabolism35. This effect could be due to end-product fermentation as the enterocytes absorb short-chain fatty acids and stimulate the growth of beneficial bacteria such as Bacillus spp. in the host intestine36.
To prevent the effects of pathogens, fish had developed a broad range of defense mechanisms19. To amplify our knowledge, immune parameters in tilapia larvae homogenates were studied. The antiprotease activity was analyzed through the % of inhibition for the trypsin activity. When comparing FOS results with the control group, significant differences were detected. Antiprotease activity stimulated by GHRP-6 showed a positive modulation, though statistically, it was insignificant concerning the control group. Lysozyme in homogenates was also measured, showing significant differences for both dietary supplementations.
A higher level of disease resistance in fish has been proven to be related to stimulating these activities17,18,19. Antiproteases are enzyme inhibitors which primarily target the internal protein and polypeptide structure of pathogens37. Consistent with our study, Sirimanapong et al. 201538 found a 60% of trypsin inhibition in P. hypophthalmus fed with the immunostimulant β-glucan. Meanwhile, lysozymes had been demonstrated to induce specific and non-specific immune responses in teleosts39. Similar to these results, dietary FOS and GHRP-6 has been reported to stimulate immune responses, such as serum lysozyme of red drum and Nile tilapia, respectively19,40. Considering these facts, our results suggest that FOS and GHRP-6 administration in diets could also be involved in better disease outcomes in fish.
Figure. 4. Nutritional supplement of FOS and GHRP-6 modulated digestive enzyme activities and immune parameters in tilapia (Oreochromis niloticus).
CONCLUSIONS
To our knowledge, this study is the first to show a significant improvement in growth parameters when feeding tilapia larvae with 0.4% FOS on diet. In addition, our results are complemented by a study of the stimulation of digestive enzyme activity and immune system parameters, in which the FOS diet also showed enhancing effects. Also, we developed these exact measurements for GHRP-6, obtaining positive novel results as well.
In conclusion, FOS and GHRP-6 dietary supplements could be exciting for commercial aquaculture to be included as additives in food. This also stands as a base for further studies in which both diets can act synergistically or whether their effects complement each other, allowing better use of food, considering the digestive enzyme activities and immune responses stimulated by the peptide.
Acknowledgments
The authors thank the colleagues Mirtha Binlloy, José Llanes and Teresita Damas from the Aquaculture Technology Development Company of Havana, Cuba, for participating in this work.
REFERENCES
1. Oecd, F. A. O. (2022). OECD-FAO Agricultural Outlook 2022-2031.
2. Jep Lokesh, Mylène Ghislain, Marine Reyrolle, Mickael Le Béchec, Thierry Pigot, et al. Prebiotics Modify Host Metabolism in Rainbow Trout (Oncorhynchus Mykiss) Fed with a Total Plant-Based Diet: Potential Implications for Microbiome-Mediated Diet Optimization. Aquaculture, 2022, 561, pp.738699. https://doi.org/10.1016/j.aquaculture.2022.738699hal-03763673
3. Maclean, N.; M.A. Rahman, F. Sohm, G. Hwang, A. Iyengar, H. Ayad, A. Smith y H. Farahmand (2002): Transgenic tilapia and tilapia genome. Gene. 295(265-277).
4. Bruzzese E, Volpicelli M, Squaglia M, Tartaglione A, Guarino A. Impact of prebiotics on human health. Dig Liver Dis. 2006;38:S283–7. https://www.sciencedirect.com/science/article/pii/S1590865807600115
5. Guarino MPL, Altomare A, Emerenziani S, Di Rosa C, Ribolsi M, Balestrieri P, et al. Mechanisms of action of prebiotics and their effects on gastrointestinal disorders in adults. Nutrients. 2020;12. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7231265/
6. Guerreiro I Couto A Machado M Castro C Pousao Ferreira P Oliva-Teles A 2016. Probiotics effect on immune and hepatic oxidative status and gut morphology of white sea bream (Diplodus sargus) Fish Shellfish Immunol 50:168-174
7. Guerreiro I, Oliva‐Teles A, Enes P. Prebiotics as functional ingredients: Focus on Mediterranean fish aquaculture. Rev Aquac. 2018;10:800–32. https://onlinelibrary.wiley.com/doi/abs/10.1111/raq.12201
8. Ringø E, Olsen RE, Gifstad T, Dalmo RA, Amlund H, Hemre GI, et al. Prebiotics in aquaculture: A review. Aquac Nutr. 2010;16:117–36. https://onlinelibrary.wiley.com/doi/10.1111/j.1365-2095.2009.00731.x
9. Jia, E., Li, Z., Xue, Y., Jiang, G., Li, X., Liu, W., & Zhang, D. (2017). Effects of dietary fructooligosaccharide on the growth, antioxidants, immunity and disease resistance of Chinese mitten crab. Aquaculture, 481, 154-161. http://dx.doi.org/10.1016/j.aquaculture.2017.08.033
10. Chen, W.W., Romano, N., Ebrahimi, M., Natrah, I., 2017. The effects of dietary fructooligosaccharide on growth, intestinal short chain fatty acids level and hepatopancreatic condition of the giant freshwater prawn (Macrobrachium rosenbergii) postlarvae. Aquaculture 469, 95–101.
11. Wu, Y., Liu, W. B., Li, H. Y., Xu, W. N., He, J. X., Li, X. F., & Jiang, G. Z. (2013). Effect of dietary supplementation of fructooligosaccharide on growth performance, body composition, intestinal enzymes activities and histology of blunt snout sea bream (Megalobrama amblycephala) fingerlings. Aquaculture Nutrition, 19, 886–894.
12. Zhang, Q., Yu, H. R., Tong, T., Tong, W. P., Dong, L. F., Xua, M. Z., & Wang, Z. C. (2014). Dietary supplementation of Bacillus subtilis and fructooligosaccharide enhances the growth and non-specific immunity of juvenile ovate pompano, Trachinotus ovatus and its disease resistance against Vibrio vulnificus. Fish & Shellfish Immunology, 38, 7–14. https://doi.org/10.1016/j.fsi.2014.02.008
13. Poolsawat, L., Li, X., Yang, H., Yang, P., Kabir Chowdhury, M. A., Yusuf, A., & Leng, X. (2020). The potentials of fructooligosaccharide on growth, feed utilization, immune and antioxidant parameters, microbial community and disease resistance of tilapia (Oreochromis niloticus× O. aureus). Aquaculture Research, 51(11), 4430-4442.
14. Martinez, R., Carpio, Y., Morales, A., Lugo, J. M., Herrera, F., Zaldívar, C., Carrillo, O., Arenal, A., Pimentel, E., & Estrada, M. P. (2016a). Oral administration of the growth hormone secretagogue-6 (GHRP-6) enhances growth and non-specific immune responses in tilapia (Oreochromis sp.). Aquaculture, 452, 304–310. https://doi.org/10.1016/j.aquaculture.2015.11.014
15. Martínez, R., Hernández, L., Gil, L., Carpio, Y., Morales, A., Herrera, F., Rodríguez-Mallón, A., Leal, Y., Blanco, A., & Estrada, M. P. (2017a). Growth hormone-releasing peptide-6 enhanced antibody titers against subunit antigens in mice (BALB/c), tilapia (Oreochromis niloticus) and African catfish (Clarias gariepinus). Vaccine, 35(42), 5722–5728. https://doi.org/10.1016/j.vaccine.2017.07.060
16. Martínez, R., Carpio, Y., Arenal, A., Lugo, J. M., Morales, R., Martín, L., Rodríguez, R. F., Acosta, J., Morales, A., Duconge, J., & Estrada, M. P. (2017b). Significant improvement of shrimp growth performance by growth hormone-releasing peptide-6 immersion treatments. Aquaculture Research, 48(9), 4632–4645. https://doi.org/10.1111/are.13286
17. Yengkhom, O., Shalini, K. S., Subramani, P. A., & Michael, R. D. (2019). Stimulation of non-specific immunity, gene expression, and disease resistance in Nile Tilapia, Oreochromis niloticus (Linnaeus, 1758), by the methanolic extract of the marine macroalga, Caulerpa scalpelliformis. Veterinary World, 12(2), 271.
18. Li, R., Yang, X., Li, Q., Ye, S., & Li, H. (2015). Enhanced immune response and resistance to edwardsiellosis following dietary chitooligosaccharide supplementation in the olive flounder (Paralichthys olivaceus). Fish and Shellfish Immunology, 47(1), 74–78. https://doi.org/10.1016/j.fsi.2015.08.019
19. Hernández L, Camacho H, Nuñez-Robainas A, Palenzuela DO, Morales A, Basabe L, Herrera F, Rodrigo O, Rodriguez-Gabilondo A, Velázquez J, Piloto S, Estrada MP, Martínez R, Growth hormone secretagogue peptide-6 enhances Oreochromicins transcription and antimicrobial activity in tilapia (Oreochromis sp.), Fish and Shellfish Immunology (2021), doi: https://doi.org/10.1016/j.fsi.2021.08.011
20. Gella, F. J., Gubern, G., Vidal, R., & Canalias, F. (1997). Determination of total and pancreatic α-amylase in human serum with 2-chloro-4-nitrophenyl-α-D-maltotrioside as substrate. Clinica Chimica Acta, 259(1-2), 147-160.
21. Gilham, D., & Lehner, R. (2005). Techniques to measure lipase and esterase activity in vitro. Methods, 36(2), 139-147.
22. Martinez, R., Ubieta, K., Herrera, F., Forellat, A., Morales, R., De la Nuez, A., Rodriguez, R., Reyes, O., Oliva, A., & Estrada, M. P. (2012). A novel GH secretagogue, A233, exhibits enhanced growth activity and innate immune system stimulation in teleosts fish. Journal of Endocrinology, 214(3), 409–419. https://doi.org/10.1530/JOE-11-0373
23. Soleimani, N., Hoseinifar, S. H., Merrifield, D. L., Barati, M., & Abadi, Z. H. (2012). Dietary supplementation of fructooligosaccharide (FOS) improves the innate immune response, stress resistance, digestive enzyme activities and growth performance of Caspian roach (Rutilus rutilus) fry. Fish & Shellfish Immunology, 32(2), 316–321. https://doi:10.1016/j.fsi.2011.11.023
24. Teitelbaum, J. E., & Walker, W. A. (2002). Nutritional impact of pre-and probiotics as protective gastrointestinal organisms. Annual review of nutrition, 22(1), 107-138.
25. Ringø, E., & Song, S. K. (2016). Application of dietary supplements (synbiotics and probiotics in combination with plant products and β‐glucans) in aquaculture. Aquaculture Nutrition, 22(1), 4-24.
26. Swanson, K., Grieshop, C., Flickinger, E., Healy, H. P., Dawson, K. A., Merchen, N. R., ... & Fahey Jr, G. C. (2002). Effects of supplemental fructooligosaccharides plus mannanoligosaccharides on immune function and ileal and fecal microbial populations in adult dogs. Archives of Animal Nutrition, 56(4), 309-318.
27. Rodríguez-Viera, L.; Martí, I.; Martínez, R.; Perera, E.; Estrada, M.P.; Mancera, J.M.; Martos-Sitcha, J.A. Feed Supplementation with the GHRP-6 Peptide, a Ghrelin Analog, Improves Feed Intake, Growth Performance and Aerobic Metabolism in the Gilthead Sea Bream Sparus aurata. Fishes 2022, 7, 31. https://doi.org/10.3390/fishes7010031
28. Lugo, J.M.; Oliva, A.; Morales, A.; Reyes, O.; Garay, H.E.; Herrera, F.; Cabrales, R.; Perez, E.; Estrada, M.P. The biological role of pituitary adenylate cyclase-activating polypeptide (PACAP) in growth and feeding behavior in juvenile fish. J. Pept. Sci. 2010, 16, 633–643
29. Martínez, R.; Morales, C.; Arenal, A.; Morales, A.; Herrera, F.; González, V.; Estrada, M.P. Growth Hormone Secretagogue (A233) Improves Growth and Changes the Tissue Fatty Acid Profile in Juvenile Tilapia (Oreochromis niloticus). Lipids 2018, 53, 429–436
30. Unniappan, S.; Lin, X.; Cervini, L.; Rivier, J.; Kaiya, H.; Kangawa, K.; Peter, R.E. Goldfish ghrelin: Molecular characterization of the complementary deoxyribonucleic acid, partial gene structure and evidence for its stimulatory role in food intake. Endocrinology 2002, 143, 4143–4146.
31. Grisdale-Helland, B., Helland, S. J., & Gatlin, D. M. (2008). The effects of dietary supplementation with mannooligosaccharide, fructooligosaccharide or galactooligosaccharide on the growth and feed utilization of Atlantic salmon (Salmo salar). Aquaculture, 283(1–4), 163–167.
32. Ai, Q., Xu, H., Mai, K., Xu, W., Wang, J., & Zhang, W. (2011). Effects of dietary supplementation of Bacillus subtilis and fructooligosaccharide on growth performance, survival, non-specific immune response and disease resistance of juvenile large yellow croaker, Larimichthys crocea. Aquaculture, 317(1-4), 155-161.
33. Ye, J. D., Wang, K., Li, F. D., & Sun, Y. Z. (2011). Single or combined effects of fructose-and mannan oligosaccharide supplements and Bacillus clausii on the growth, feed utilization, body composition, digestive enzyme activity, innate immune response and lipid metabolism of the Japanese flounder Paralichthys olivaceus. Aquaculture Nutrition, 17, 902–911.
https://doi.org/10.1111/j.1365-2095.2011.00863.x
34. Furne, M., Hidalgo, M. C., Lopez, A., Garcia-Gallego, M., Morales, A. E., Domezain, A., ... & Sanz, A. (2005). Digestive enzyme activities in Adriatic sturgeon Acipenser saccharin and rainbow trout Oncorhynchus mykiss. A comparative study. Aquaculture, 250(1-2), 391-398.
35. Ashraf, M. Abd El-latif, Eman, A. Abd El-Gawad and Mahmoud A. Emam. Effect of dietary fructooligosaccharide supplementation on feed utilization and growth performance of Nile tilapia (Oreochromis niloticus) fingerlings. Egy. J. Aquac., Vol 5, No. (3):1-16 (2015) ISSN: 2090-7877
36. Mahious, A. S., Gatesoupe, F. J., Hervi, M., Metailler, R., & Ollevier, F. (2006). Effect of dietary inulin and oligosaccharides as prebiotics for weaning turbot, Psetta maxima (Linnaeus, C. 1758). Aquaculture International, 14, 219–229. https://doi.org/10.1007/s10499-005-9003-4
37. Adikesavalu, H., Hoque, F., & Abraham, T. (2020). Immune effector activities and clinical biochemistry of normal pangas catfish Pangasius pangasius (Hamilton, 1822). International Aquatic Research, 12(4), 301.
38. Sirimanapong W, Adams A, Ooi EL, Green DM, Nguyen DK, Browdy CL, Collet B, Thompson KD, The effects of feeding immunostimulant β-glucan on the immune response of Pangasianodon hypophthalmus, Fish and Shellfish Immunology (2015). https://doi:10.1016/j.fsi.2015.04.025
39. Shakoori, M., Hoseinifar, S. H., Paknejad, H., Jafari, V., Safari, R., Van Doan, H., & Mozanzadeh, M. T. (2019). Enrichment of rainbow trout (Oncorhynchus mykiss) fingerlings diet with microbial lysozyme: Effects on growth performance, serum and skin mucus immune parameters. Fish & shellfish immunology, 86, 480-485.
40. Buentello JA, Neill WH, Gatlin III DM. Effects of dietary prebiotics on the growth, feed efficiency and non-specific immunity of juvenile red drum Sciaenops ocellatus fed soybean-based diets. Aquaculture Research 2010;41:411e8.
Received: 28 May 2023/ Accepted: 15 July 2023 / Published:15 September 2023
Citation: Morales A, Moro D F, Rodriguez A, Hernández L, Comellas A, Herrera F, Gonzalez O, Pérez Cruz E, Estrada M P, Martinez R. Nutritional supplement of FOS enhances growth and immune system in tilapia larvae (Oreochromis niloticus), Revis Bionatura 2023;8 (3) 22. http://dx.doi.org/10.21931/RB/2023.08.03.22