2023.08.01.19
Files > Volume 8 > Vol 8 No 1 2023
Microorganisms isolated from seabirds feathers for mercury bioremediation

Lorena Monserrate-Maggi 1 , Lizette Serrano-Mena 1
, Lizette Serrano-Mena 1 , Louise Delahaye 2, Paola Calle 3*
, Louise Delahaye 2, Paola Calle 3* , Omar Alvarado-Cadena3, Omar Ruiz-Barzola 3,4
, Omar Alvarado-Cadena3, Omar Ruiz-Barzola 3,4 , Juan Manuel Cevallos-Cevallos 1,3
, Juan Manuel Cevallos-Cevallos 1,3 *
*
1 Escuela Superior Politécnica del Litoral, ESPOL, Centro de Investigaciones Biotecnológicas del Ecuador, CIBE, Campus Gustavo Galindo Km 30.5 Vía Perimetral, P.O Box 09-01-5863, Guayaquil, Ecuador; [email protected] . [email protected] . [email protected] .
2 VIVES University CollegePost-graduate International Cooperation North-South, Kortrijk, Belgium; [email protected]; [email protected]
3 Escuela Superior Politécnica del Litoral, ESPOL, Facultad de Ciencias de la Vida, FCV, Campus Gustavo Galindo Km 30.5 Vía Perimetral, P.O Box 09-01-5863, Guayaquil, Ecuador; [email protected]; [email protected] .
4 Universidad de Salamanca, Departamento de Estadística; Campus Miguel de Unamuno. 37007, Salamanca -España; [email protected]
* Correspondence: [email protected]; P,C. [email protected]; J,C-C.
Available from. http://dx.doi.org/10.21931/RB/2023.08.01.19
ABSTRACT
Environmental pollution caused by mercury has received increasing attention in recent years. Several studies have warned of the high rates of biomagnification in superior levels of marine food networks affecting seabirds. Although seabird feathers are reported as bioindicators of mercury, the possibility of using the microbiota associated with them for the bioremediation of this metal has not been considered. Despite the potential of the seabird feather microbiota, the cultivable microorganisms from this sample matrix have not been identified. In this study, we isolated and identified the organisms in the feathers from three types of seabirds, two species of penguins (Pygoscelis antartica and Pygoscelis papua) and the brown skua bird (Catharacta lonnbergi) through poisoned media a final concentration of 10 mg / L Hg2+ in the culture medium for the microbial consortia. Yeast isolates belonged to the genus Debaryomyces, Meyerozyma, Papiliotrema, and Rhodotorula, and fungi genera Leiotrametes, Penicillium, Pseudogymnoascus, and Cladosporium were identified. Adult bird feathers with high mercury concentrations can serve as a matrix to isolate microorganisms capable of removing mercury.
Keywords: Antarctica, bioremediation, feathers, mercury, microorganisms
INTRODUCTION
Mercury (Hg) is among the most severe pollutants due to its accumulation in food chains, resulting in risks to human, animal, and environmental health 1–4. The atmospheric transport of this metal affects the most remote and cleanest areas of the planet, such as the polar zones, reaching even higher levels of deposition than in other parts of the world 5–8, thus affecting the aquatic ecosystems of Antarctica 9,10.
The elemental mercury (Hg0) and ionic mercury (Hg2+) that reach Antarctica fall on sediment and water bodies, while some elemental mercury remains dissolved in the water column. Another part of mercury is transformed by microorganisms, through the biomethylation process, to a more toxic organometallic compound, the methylmercury (CH3Hg+), which will be bioaccumulated and biomagnified along the marine trophic chain 11. With a trophic magnification factor (TMF) of 4 to 8 for each step of the trophic level 12,13, the amount of CH3Hg+ in predatory species can be up to 100 times higher than their primary food source so that birds, among other species, are the most exposed in the marine ecosystems of Antarctica 9. In addition to its high position in the food chain, the bioaccumulation of Hg in birds is favored by factors such as its wide distribution, population variety, long life cycles, and its type of diet 14,15.
Although in Antarctica there is no industrial development that contributes to mercury emissions 4,6,16, its proximity to the southern hemispheres, the tourism, the pollution from logistics activities of scientific stations 17–20 as well as contamination of natural origin from volcanic activity 10,21 contribute to the increase of Hg in predatory seabirds from different locations in this continent 14,22.
Complex communities of microorganisms are found in birds' feathers whose composition can be influenced by exposure to heavy metals 23. It is well known that organisms living in contaminated or toxic conditions have developed different mechanisms to adapt to high levels of various forms of mercury present in the environment and can be used for bioremediation or mitigation of the contaminant 24–26. Bioremediation is an option that uses those strategies that microorganisms have developed to deal with Hg, with exceptional advantages that include high efficiency, low cost and environmentally friendly 27. Therefore, it is essential to identify the microorganisms living in high-mercury environments 28–30.
Current research primarily focuses on the ability of Antarctic seabird feathers to act as bioindicators of Hg 31–33 or in the isolation of microorganisms from soils and water contaminated with Hg 24,25,27–30. However, the microbiota associated with bird feathers having high mercury levels has not been fully described.
Therefore, the objective of this study was to identify the culturable microorganisms from the feathers of three Antarctic seabirds known to biomagnify mercury, including the geentoo penguins Pygoscelis papua and chinstrap Pygoscelis antarctica and the skuas brown Catharacta lonnbergi, which inhabit the surroundings of the Pedro Vicente Maldonado Scientific Station in Antarctica, as a first step that can aid further mercury bioremediation studies.
MATERIALS AND METHODS
Sample collection
The present investigation corresponds to an exploratory study using a purposive (judgmental) sampling method. The seabird feather samples were collected during the scientific expeditions to Antarctica carried out by the Instituto Antártico Ecuatoriano (INAE) during the summer of 2013 and 2014. The sampling was carried out in the surrounding areas of the Ecuadorian Scientific Station Pedro Vicente Maldonado (PEVIMA), located in the South Shetland archipelago of the Antarctic Peninsula. The islands evaluated were Barrientos (n = 2 sites), Dee (n = 1 site) and Greenwich (n = 2 sites). Figure 1 shows the sampling sites.

Figure 1. The geographical location of the sampling sites.
The molting feathers were collected using a non-invasive method and following the guidelines of the Antarctic Treaty (1959), in which animal welfare is preserved and the capture of living individuals is avoided (34). Therefore, fallen feathers were randomly collected in nests and colonies of three bird species: P. antarctica and P. papua, corresponding to chicks and adults with feathers lengths measured between 3-6 cm, while C. lonnbergi, compared to juveniles and adults with feathers lengths measured between 15-30 cm. The description of the samples is shown in table 1.
The samples were collected in Ziploc bags and delivered to the PEVIMA station laboratory, where they were rinsed with deionized water, dried at room temperature, wrapped in aluminum foil, and kept in the freezer (-20oC) until they were analyzed at the Centro de Investigaciones Biotecnológicas del Ecuador (CIBE-ESPOL) in Guayaquil, Ecuador.
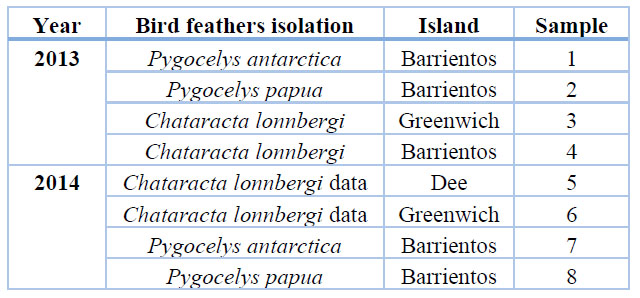
Table 1. Origin of the collection of feathers by years.
Isolation of microorganisms from feathers
The feathers were subjected to an individual cleaning process in which the barbs exposed to external conditions and/or feather age, which could influence the interpretation of the results, were removed 35,36. The rachis was cleaned with hypochlorite solution (30%) for 30 seconds and immersed in 99% and 70% ethanol for 30 seconds each. Finally, they were rinsed with plenty of ultrapure water. The rachis of the feathers was then ground under liquid nitrogen (LN2) in a porcelain mortar and collected in 15-ml falcon tubes to enrich microorganisms in the feathers.
For the enrichment process of the samples, 1 gr of the crushed sample was weighed and placed in a 15ml falcon tube with 9ml of liquid culture medium. The culture media used were Peptone water (AP, Oxoid, Thermo Scientific, USA), Luria Bertani (LB, Oxoid, Thermo Scientific, USA), and Potato Dextrose Broth (PDB, Oxoid, Thermo Scientific, USA) that were previously autoclaved at 121 °C for 25 minutes. Then, the samples were incubated at 10 °C for seven days with constant shaking at 110 rpm (Innova 44R, New Brunswick, USA).
Then, to determine the tolerance of microorganisms to Hg, the reference standard of inorganic mercury, Hg2 + (HACH, Germany), was added, using aseptic techniques, at a concentration 10 times higher than that reported in certain feathers of Antarctic birds 31 and taking into account the toxicity threshold for adverse effects in seabird feathers of between 5-30 mg / L as reported by some authors 14,37. For this reason, a final concentration of 10 mg / L Hg2+ in the culture medium was used for the microbial consortia and 5mg / L Hg2+ for the isolates cultured from the microbial consortia. The three poisoned media without samples were used as a blank, and each treatment was performed in duplicate.
Isolation and molecular identification of microorganisms present in the consortia feathers
Seven days after adding Hg to the microbial cultures of bird feather consortia, 100ul of the culture were taken and dispersed in Petri dishes previously prepared with 57.5 g / L of Potato Dextrose Agar medium (PDA, Oxoid, Thermo Scientific, USA) plus 10 mg / L Hg2+, and incubated at 10°C for isolation of cultivable microorganisms. After this, the obtained strains were separated into yeasts and fungi according to their macro and micromorphology.
The DNA extraction from isolates was carried out using a rapid fungal DNA extraction protocol according to Cenis (1992) 38. Molecular identification was performed by PCR amplification and sequencing of the internal transcribed spacer regions (ITS1, 5.8S, and ITS2) using ITS1 (TCCGTAGGTGAACCTGCGG) and ITS4 (TCCTCCGCTTATTGATATGC) primers, the samples that did not amplify, a PCR of nesting using primers ITS3 (GCTTCGATGAAGAACGCAGC) and ITS4. The master mix for both PCRs was: 1X PCR buffer, 0.2 mM dNTP, 1.5 mM MgCl2, 5 U / µL Taq polymerase (ThermoFisher, USA), 0.4 µM of each primer, and the DNA concentration comprised 15-20ng / ul. The program in the thermal cycler (Eppendorf, Mastercycler Nexus GSX1-6345, Germany) for the first set of primers consisted of initial heating of 1 minute at 94 °C, followed by 30 cycles of 1 minute at 94 °C, 1 min at 55 °C, 1 min at 68 °C, and a final extension for 3 min at 68 °C. For the second set of primers, the PCR conditions were: 6 min at 95 ° C, 30 cycles from 0:30 min to 95 ° C, 0:30 min to 55 ° C, 0:30 min to 70 ° C, followed by a final extension of 0:30 min at 72 ° C. Amplification was verified by electrophoresis of a 1.5% agarose gel in 1X TAE solution (Tris base, boric acid, and 0.5M EDTA, pH 8.0), loading 5 µL of PCR product with 1 µL of loading dye (Loading dye, Promega, USA) at 100 volts for 30 minutes. The size of each DNA fragment was estimated using a 100 bp DNA marker (cat. 15628050, Invitrogen ™). Gel images were analyzed using the Gel Doc XR Imager program (Bio-Rad, Philadelphia, PA).
The obtained PCR products were sequenced by Sanger at Macrogen, Inc., an external laboratory in South Korea, according to Genetic Resource Access Contract No MAE-DNB-CM-2017-0059, material transfer agreement MAE-DNB-CM-2017-0059-000-ATM-0001, and sample export authorization No 074-17-EXP-IC-FAU-DNB/MA granted by the Ministry of the Environment of Ecuador.
The chromatograms of the DNA sequences of the different isolates were visualized and edited manually using the program Finch TV version 1.4.0 (Geospiza Inc.) and then compared with the database of the National Center for Biotechnological Information (NCBI) using BLAST. The sequences were deposited in the GenBank public database (http://www.ncbi.nlm.nih.gov/GenBank), and the isolates collection resides in the Microorganism Culture Collection of CIBE at Escuela Superior Politécnica del Litoral (http://www.wfcc.info/ccinfo/index.php/collection/by_id/1151/).
RESULTS AND DISCUSSION
Seabirds' bioaccumulation mercury has been reported from different locations in the Antarctic and including in petrel feathers Pagodroma nivea 0.54 ± 0.18 μg g−1 dry wt 18, antarctic petrel Thalassoica antarctica 2.71 ± 0.25 mg g-1 dw 39; Gentoo penguins Pygoscelis papua 1.83 ± 0.80 ug g−1 dw 31,40–42, Chinstrap penguins Pygoscelis antarctica 1.53 ± 0.08 ug g−1 dw 31,40,41, Pygoscelis adeliae 0.82 to 1.40 ± 0.13 μg g−1 dw 18,40,41, Aptenodytes forsten 0.98 ± 0.2 μg g−1 dw; skuas Catharacta maccormicki 2.91 ± 1.93 µg g -1 dw 18, Catharacta lonnbergi 2.86 ± 2.60 ug g−1 dw 31 and gull Larus dominicanus 426.6 ng g−1 9.
In this study, isolated species of seabirds used in this study corresponding to the following yeast genera were identified: Debaryomyces, Meyerozyma, Papiliotrema, and Rhodotorula also fungi genera: Leiotrametes, Penicillium, Pseudogymnoascus, and Cladosporium. The most abundant species in the bird consortiums correspond to yeasts of the genus Debaromyces followed by the fungi genus Pseudogymnoascus and Penicillum. Table 2 and table S1. Some of the yeasts belonging to the Rhodotorula and family Saccharomycetaceae and fungi between these Cladosporium and Penicillium have been reported with efficient accumulation strategies and biovolatilization of mercury regardless of their origin. They are considered suitable for application in remedial technology 43–45.

Table 2. Molecular identification according to ITS region of the cultivable isolates.
Regarding the efficiency of the culture media for metal removal, they were AP and PDB, and the fact that only yeasts and fungi have been isolated may be due to these culture media being a broad spectrum range favoring the growth of these groups 46.
On the other hand, the differences in the level of resistance to metals among genera and strains depend on different growth requirements (such as temperature, pH, and nutrients), biological function 47, or pressure origin 43.
One of the mechanisms responsible for removing mercury in the medium from cultivable isolated may be due to the biosorption capacity of the fungal cell wall, which contains polysaccharides with reactive functional groups, amino, carboxyl, and phosphate. Of these, it is known that the carboxyl and phosphate groups carry negative charges that allow fungal cell wall components to be highly metal ion-retaining 48. The peptide links of nitrogen and oxygen could be accompanied by the displacement of protons, depending on the pH, which also favors the removal of the metal 49,50. The number of available binding sites determines metal biosorption 51. On the other hand, fungal mycelium secretes many extracellular enzymes and acids that decompose metals and has a huge potential for degrading contaminants 45,52,53.
Urík et al. (2014) 43 indicated fungal mercury uptake increases linearly with increased initial media mercury concentration until a threshold concentration near 8.2 mg L−1. When the amount of mercury remaining in media with higher than threshold concentration decreased by 75 % or more, sorption via mercury immobilization on the fungal cell wall and bioaccumulation in the intracellular compartments play insignificant roles in mercury resistance strategy; hence the fungal necessity to trigger other detoxification mechanisms confirm that biovolatilization is the main mechanism of detoxification of mercury by fungal strains. Also, some authors confirm that biovolatilization is the primary mechanism of the detoxification of mercury by fungal strains43).
Other fungi strains isolated from soil samples like Aspergillus niger removed more than 90% and proved an excellent mercury absorber. Aspergillus flavus strain and Cladosporium can eliminate more than 90% of 10 mg L−1 of initial mercury concentration in static culture for 7 days and have been reported with biovolatilization efficiency rendering them the most suitable for application in remedial technology 43,54,55.
The precise fungal mercury volatilization mechanism is not currently elucidated, but it most likely involves some intra or extracellular reducing factor and/or methylation agent when considering mercury volatilization in dimethyl form 56,57. However, it should not be ruled out that another mechanism of action of the cultivable isolates proposed by Kelly et al. (2006) where mercury deposition as HgS in microfungi dominates at low mercury concentrations 58.
Findings focused on bioremediation, comparing the use of consortia (multiple or heterogeneous systems) with pure isolates (homogeneous systems), describe the advantages of living in the community. Many factors can influence passive and active mechanisms in the removal of metals, as well as considering the relationship with the use of carbon sources and biodegradation processes59, and they can withstand higher concentrations of heavy metals. The use of these represents a closer approximation to what occurs in nature. This also allows the development of experimental model systems, which can explain the lag between the boiadsorption of pure cultures in situ 60. While pure isolates can resist lower concentrations of mercury compared to consortia, and few mechanisms of action on metal could be focused on, like bioaccumulation, biosorption, biopreciping, and/ or biovolitizion 45,52,59.
CONCLUSIONS
Microorganisms isolated from bird feathers that biomagnify Hg in Antarctica are yeast genera identified: Debaryomyces, Meyerozyma, Papiliotrema, and Rhodotorula also, fungi genus: Leiotrametes, Penicillium, Pseudogymnoascus, and Cladosporium. Of these, yeasts belonging to the genus Rhodotorula and family Saccharomycetaceae and fungi between these Cladosporium and Penicillium have been reported with efficient strategies of accumulation and biovolatilization of mercury and are considered suitable for application in remedial technology according to the reported bibliography.
This study opens the opportunity for bioprospecting microorganisms isolated from other matrices, not mercury-contaminated water and soil, but bird feathers that biomagnified this metal. However, is a need to evaluate in time and elucidate the mechanisms used for these microorganisms in mercury removal and include other factors such as growth requirements between these pH and temperature, tolerance indices to major concentrations, and evaluation of their potential as adsorbents a low-cost and environmentally friendly. Therefore, the bioremediation of mercury from microorganisms isolated from bird feathers is still a developing technology.
Supplementary Materials: Table S1. Cultivable isolates with source and Genbank accession number.
Author Contributions: "Conceptualization, Cevallos-Cevallos.J., Calle.P. and Monserrate-Maggi.L.; methodology, Cevallos-Cevallos.J., Monserrate-Maggi.L.,Delahaye.L., Alvarado-Cadena. O. and Ruiz-Barzola.O.; formal analysis, Ruiz-Barzola.O. and Serrano-Mena. L.; investigation, Cevallos-Cevallos.J., Calle.P., Monserrate-Maggi.L., Delahaye.L.; resources, Calle.P., Alvarado-Cadena. O., Ruiz-Barzola.O. and Serrano-Mena. L.; data curation, Serrano-Mena. L.; writing—original draft preparation, Cevallos-Cellos. J., Monserrate-Maggi.L. and Serrano-Mena. L.; writing—review and editing, Monserrate-Maggi.L., Cevallos-Cevallos.J. and Calle.P.; visualization, Monserrate-Maggi.L., Cevallos-Cevallos.J. and Calle.P.; supervision, Calle.P. and Cevallos-Cevallos.J.; project administration, Cevallos-Cevallos.J. All authors have read and agreed to the published version of the manuscript."
Funding: "This research was funded by Instituto Antártico Ecuatoriano (INAE), Secretaría de Educación Superior, Ciencia, Tecnología e Innovación (SENESCYT) and Escuela Superior Politécnica del Litoral financed"
Data Availability Statement: The partial DNA sequences obtained in this study were deposited in the GenBank database. Accession numbers for each of the isolates are given in Table S1.
Conflicts of Interest: "The authors declare no conflict of interest."
REFERENCES
1. Macdonald RW, Harner T, Fyfe J. Recent climate change in the Arctic and its impact on contaminant pathways and interpretation of temporal trend data. Sci Total Environ. 2005;342(1–3):5–86.
2. Mason RP, Choi AL, Fitzgerald WF, Hammerschmidt CR, Lamborg CH, Soerensen AL, et al. Mercury biogeochemical cycling in the ocean and policy implications. Environ Res [Internet]. 2012;119:101–17. Available from: http://dx.doi.org/10.1016/j.envres.2012.03.013
3. Scheuhammer AM, Meyer MW, Sandheinrich MB, Murray MW. Effects of environmental methylmercury on the health of wild birds, mammals, and fish. Ambio. 2007;36(1):12–8.
4. Weinberg J. Introducción a la contaminación por Mercurio para las ONG [Internet]. Red Internacional de Eliminación de los Contaminantes Orgánicos Persistentes, (IPEN), editors. 2014. Available from: https://ipen.org/sites/default/files/documents/ipen_mercury_booklet-es.pdf
5. Ebinghaus R, Kock HH, Temme C, Einax JW, Löwe AG, Richter A, et al. Antarctic springtime depletion of atmospheric mercury. Environ Sci Technol. 2002;36(6):1238–44.
6. Gilmour C, Riedel G. Biogeochemistry of Trace Metals and Mettaloids. In: INORGANIC CHEMICALS: CYCLES AND DYNAMICS. Elsevier Inc.; 2009. p. 1–6.
7. Hylander LD, Goodsite ME. Environmental costs of mercury pollution. Sci Total Environ. 2006;368(1):352–70.
8. Nerentorp Mastromonaco M, Gårdfeldt K, Jourdain B, Abrahamsson K, Granfors A, Ahnoff M, et al. Antarctic winter mercury and ozone depletion events over sea ice. Atmos Environ. 2016;129:125–32.
9. Dos Santos IR, Silva-Filho EV, Schaefer C, Maria Sella S, Silva CA, Gomes V, et al. Baseline mercury and zinc concentrations in terrestrial and coastal organisms of Admiralty Bay, Antarctica. Environ Pollut. 2006;140(2):304–11.
10. Mão de Ferro A, Mota AM, Canário J. Pathways and speciation of mercury in the environmental compartments of Deception Island, Antarctica. Chemosphere. 2014;95:227–33.
11. Barkay T, Poulain AJ. Mercury (micro)biogeochemistry in polar environments. FEMS Microbiol Ecol. 2007;59(2):232–41.
12. Kidd K, Clayden M, Jardine T. Part Iv Bioaccumulation , Toxicity , and Metallomics Chapter 14 Bioaccumulation and Biomagnification of Mercury Through Food Webs. Environ Chem Toxicol Mercur. 2012;
13. Lavoie R, Jardine TD, Chumchall MM, Kidd KA, Campbell LM. Biomagnification rate of mercury in aquatic food webs: a world-wide meta-analysis. 2013;
14. Burger J, Gochfeld M. Marine Birds as Sentinels of Environmental Pollution. Ecohealth. 2004;263–74.
15. Taniguchi S, Montone RC, Bícego MC, Colabuono FI, Weber RR, Sericano JL. Chlorinated pesticides, polychlorinated biphenyls and polycyclic aromatic hydrocarbons in the fat tissue of seabirds from King George Island, Antarctica. Mar Pollut Bull [Internet]. 2009;58(1):129–33. Available from: http://dx.doi.org/10.1016/j.marpolbul.2008.09.026
16. Krabbenhoft DP, Sunderland EM. Global change and mercury. Science (80- ). 2013;341(6153):1457–8.
17. Amaro E, Padeiro A, Mão de Ferro A, Mota AM, Leppe M, Verkulich S, et al. Assessing trace element contamination in Fildes Peninsula (King George Island) and Ardley Island, Antarctic. Mar Pollut Bull [Internet]. 2015;97(1–2):523–7. Available from: http://dx.doi.org/10.1016/j.marpolbul.2015.05.018
18. Bargagli R, Monaci F, Sanchez-Hernandez JC, Cateni D. Biomagnification of mercury in an Antarctic marine coastal food web. Mar Ecol Prog Ser. 1998;169(June 2014):65–76.
19. Jerez S, Motas M, José M, Valera F, Javier J, Barbosa A. Concentration of trace elements in feathers of three Antarctic penguins : Geographical and interspeci fi c differences. Environ Pollut [Internet]. 2011;159(10):2412–9. Available from: http://dx.doi.org/10.1016/j.envpol.2011.06.036
20. Santos IR, Silva-Filho E V., Schaefer CEGR, Albuquerque-Filho MR, Campos LS. Heavy metal contamination in coastal sediments and soils near the Brazilian Antarctic Station, King George Island. Mar Pollut Bull. 2005;50(2):185–94.
21. Rey J, Somoza L, Martínez-Frías J. Tectonic, volcanic, and hydrothermal event sequence on Deception Island (Antarctica). Geo-Marine Lett. 1995;15(1):1–8.
22. Blévin P, Carravieri A, Jaeger A, Chastel O, Bustamante P, Cherel Y. Wide Range of Mercury Contamination in Chicks of Southern Ocean Seabirds. PLoS One. 2013;8(1).
23. Chatelain M, Frantz A, Gasparini J, Leclaire S. Experimental exposure to trace metals affects plumage bacterial community in the feral pigeon. J Avian Biol. 2016;47(4):521–9.
24. García-Sánchez M, Száková J. Biological Remediation of Mercury-Polluted Environments. Plant Met Interact Emerg Remediat Tech. 2015;311–34.
25. Gururajan K, Belur PD. Screening and selection of indigenous metal tolerant fungal isolates for heavy metal removal. Environ Technol Innov [Internet]. 2018;9:91–9. Available from: https://doi.org/10.1016/j.eti.2017.11.001
26. Matsui K, Endo G. Mercury bioremediation by mercury resistance transposon-mediated in situ molecular breeding. Appl Microbiol Biotechnol. 2018;102(7):3037–48.
27. Yin K, Wang Q, Lv M, Chen L. Microorganism remediation strategies towards heavy metals. Chem Eng J [Internet]. 2019;360(August 2018):1553–63. Available from: https://doi.org/10.1016/j.cej.2018.10.226
28. Liang X, Gadd GM. Metal and metalloid biorecovery using fungi. Microb Biotechnol. 2017;10(5):1199–205.
29. McCarthy D, Edwards GC, Gustin MS, Care A, Miller MB, Sunna A. An innovative approach to bioremediation of mercury contaminated soils from industrial mining operations. Chemosphere. 2017;184(December 2018):694–9.
30. Mishra A, Malik A. Recent advances in microbial metal bioaccumulation. Crit Rev Environ Sci Technol. 2013;43(11):1162–222.
31. Calle P, Alvarado O, Monserrate L, Cevallos J et al. Mercury accumulation in sediments and seabird feathers from the Antarctic Peninsula. Mar Pollut Bull [Internet]. 2015;91(2):410–7. Available from: http://dx.doi.org/10.1016/j.marpolbul.2014.10.009
32. Becker PH, Goutner V, Ryan PG, González-Solís J. Feather mercury concentrations in Southern Ocean seabirds: Variation by species, site and time. Environ Pollut. 2016;216:253–63.
33. Carravieri A, Cherel Y, Blévin P, Brault-Favrou M, Chastel O, Bustamante P. Mercury exposure in a large subantarctic avian community. Environ Pollut [Internet]. 2014;190:51–7. Available from: http://dx.doi.org/10.1016/j.envpol.2014.03.017
34. Metcheva R, Yurukova L, Teodorova S, Nikolova E. The penguin feathers as bioindicator of Antarctica environmental state. Sci Total Environ. 2006;362(1–3):259–65.
35. Dauwe T, Bervoets L, Pinxten R, Blust R, Eens M. Variation of heavy metals within and among feathers of birds of prey: Effects of molt and external contamination. Environ Pollut. 2003;124(3):429–36.
36. Goede AA, de Bruin M. The use of bird feather parts as a monitor for metal pollution. Environ Pollution Ser B, Chem Phys. 1984;8(4):281–98.
37. Polito MJ, Brasso RL, Trivelpiece WZ, Karnovsky N, Patterson WP, Emslie SD. Differing foraging strategies influence mercury (Hg) exposure in an Antarctic penguin community. Environ Pollut. 2016;218:196–206.
38. Cenis JL. Rapid extraction of fungal DNA for PCR amplification. Nucleic Acids Res. 1992;20(9):2380.
39. Carravieri A, Fort J, Tarroux A, Cherel Y, Love OP, Prieur S, et al. Mercury exposure and short-term consequences on physiology and reproduction in Antarctic petrels. Environ Pollut. 2018;237:824–31.
40. Álvarez-Varas R, Morales-Moraga D, González-Acuña D, Klarian SA, Vianna JA. Mercury Exposure in Humboldt (Spheniscus humboldti) and Chinstrap (Pygoscelis antarcticus) Penguins Throughout the Chilean Coast and Antarctica. Arch Environ Contam Toxicol [Internet]. 2018;75(1):75–86. Available from: https://doi.org/10.1007/s00244-018-0529-7
41. Brasso RL, Chiaradia A, Polito MJ, Raya Rey A, Emslie SD. A comprehensive assessment of mercury exposure in penguin populations throughout the Southern Hemisphere: Using trophic calculations to identify sources of population-level variation. Mar Pollut Bull [Internet]. 2015;97(1–2):408–18. Available from: http://dx.doi.org/10.1016/j.marpolbul.2015.05.059
42. Chiang G, Kidd KA, Díaz-Jaramillo M, Espejo W, Bahamonde P, O'Driscoll NJ, et al. Methylmercury biomagnification in coastal aquatic food webs from western Patagonia and western Antarctic Peninsula. Chemosphere [Internet]. 2021;262(128360). Available from: https://doi.org/10.1016/j.chemosphere.2020.128360
43. Urík M, Hlodák M, Mikušová P, Matúš P. Potential of microscopic fungi isolated from mercury contaminated soils to accumulate and volatilize mercury(II). Water Air Soil Pollut. 2014;225(2219).
44. Kordialik-Bogacka E. Surface properties of yeast cells during heavy metal biosorption. Cent Eur J Chem. 2011;9(2):348–51.
45. Mustapha MU, Halimoon N. Microorganisms and Biosorption of Heavy Metals in the Environment: A Review Paper. J Microb Biochem Technol [Internet]. 2015;07(05):253–6. Available from: https://www.omicsonline.org/open-access/microorganisms-and-biosorption-of-heavy-metals-in-the-environment-a-review-paper-1948-5948-1000219.php?aid=60592
46. American Type Culture Collection A. Mycology Culture Guide Table of Contents [Internet]. Manassas; 2013. 33 p. Available from: www.atcc.org
47. Ezzouhri L, Castro E, Moya M, Espinola F, Lairini K. Heavy metal tolerance of filamentous fungi isolated from polluted sites in Tangier, Morocco. African J Microbiol Res. 2009;3(2):35–48.
48. Say R, Yilmaz N, Denizli A. Biosorption of cadmium, lead, mercury, and arsenic ions by the fungus Penicillium purpurogenum. Sep Sci Technol. 2003;38(9):2039–53.
49. Gow NAR, Latge J, Munro CA, De Groot PWJ, Hellingwerf KJ, Klis FM, et al. Cell Wall Architecture in Yeast : New Structure and New Challenges MINIREVIEW Cell Wall Architecture in Yeast : New Structure and New Challenges †. Yeast. 2003;9(3):3341–54.
50. Ozsoy HD. Biosorptive removal of Hg(II) ions by Rhizopus oligosporus produced from corn-processing wastewater. African J Biotechnol. 2010;9(51):8791–9.
51. Rezaee A, Derayat J, Mortazavi SB, Yamini Y, Jafarzadeh MT. Removal of Mercury from chlor-alkali Industry Wastewater using Acetobacter xylinum Cellulose. Am J Environ Sci. 2005;1(2):102–5.
52. Gupta A, Joia J. Microbes as Potential Tool for Remediation of Heavy Metals: A Review. J Microb Biochem Technol. 2016;8(4):364–72.
53. Veglio F, Beolchini F. Removal of metals by biosorption: A review. Hydrometallurgy. 1997;44(3):301–16.
54. Joo J-H, Hussein KA. Heavy Metal Tolerance of Fungi Isolated from Contaminated Soil. Korean J Soil Sci Fertil. 2012;45(4):565–71.
55. Oladipo OG, Awotoye OO, Olayinka A, Bezuidenhout CC, Maboeta MS. Heavy metal tolerance traits of filamentous fungi isolated from gold and gemstone mining sites. Brazilian J Microbiol [Internet]. 2018;49(1):29–37. Available from: http://dx.doi.org/10.1016/j.bjm.2017.06.003
56. Jiménez-Moreno M, Perrot V, Epov VN, Monperrus M, Amouroux D. Chemical kinetic isotope fractionation of mercury during abiotic methylation of Hg(II) by methylcobalamin in aqueous chloride media. Chem Geol [Internet]. 2013;336:26–36. Available from: http://dx.doi.org/10.1016/j.chemgeo.2012.08.029
57. Yannai S, Berdicevsky I, Duek L. Transformations of inorganic mercury by Candida albicans and Saccharomyces cerevisiae. Appl Environ Microbiol. 1991;57(1):245–7.
58. Kelly DJA, Budd K, Lefebvre DD. The biotransformation of mercury in pH-stat cultures of microfungi. Can J Bot. 2006;84(2):254–60.
59. Sprocati AR, Alisi C, Segre L, Tasso F, Galletti M, Cremisini C. Investigating heavy metal resistance, bioaccumulation and metabolic profile of a metallophile microbial consortium native to an abandoned mine. Sci Total Environ. 2006;366(2–3):649–58.
60. Ledin M. Accumulation of metals by microorganisms - processes and importance for soil systems. Earth Sci Rev. 2000;51(1–4):1–31.
Received: September 26, 2022 / Accepted: October 15, 2022 / Published:15 February 2023
Citation: Monserrate-Maggi, L.; Serrano-Mena, L.; Delahaye, L.; Calle, P.; Alvarado-Cadena, O.; Ruiz,O.; Cevallos-Cevallos, J., Microorganisms isolated from seabirds feathers for mercury bioremediation. Revis Bionatura 2023;8 (1)19. http://dx.doi.org/10.21931/RB/2023.08.01.19