2023.08.01.20
Files > Volume 8 > Vol 8 No 1 2023
Genetic improvement in Musa through modern biotechnological methods

Villao, L. 1 , Chávez, T. 1
, Chávez, T. 1 , Pacheco, R. 1
, Pacheco, R. 1 , Sánchez, E. 1,2
, Sánchez, E. 1,2 , Bonilla J. 1,2
, Bonilla J. 1,2 , Santos, E. 1,2
, Santos, E. 1,2
1 Escuela Superior Politécnica del Litoral, ESPOL, Biotechnological Research Center of Ecuador, Gustavo Galindo Campus Km. 30.5 Vía Perimetral, PO Box 09-01-5863, Guayaquil, Ecuador.
2 Escuela Superior Politécnica del Litoral, ESPOL, Faculty of Life Sciences, Gustavo Galindo Campus Km. 30.5 Vía Perimetral, PO Box 09-01-5863, Guayaquil, Ecuador.
Corresponding autor. Email: [email protected]
Available from. http://dx.doi.org/10.21931/RB/2023.08.01.20
ABSTRACT
Bananas, one of the most valued fruits worldwide, are produced in more than 135 countries in the tropics and subtropics for local consumption and export due to their tremendous nutritional value and ease of access.
The genetic improvement of commercial crops is a crucial strategy for managing pests or other diseases and abiotic stress factors. Although conventional breeding has developed new hybrids with highly productive or agronomic performance characteristics, in some banana cultivars, due to the high level of sterility, the traditional breeding strategy is hampered. Therefore, modern biotechniques have been developed in a banana for genetic improvement. In vitro, culture techniques have been a basis for crop micropropagation for elite banana varieties and the generation of methods for genetic modification. This review includes topics of great interest for improving bananas and their products worldwide, from their origins to the different improvement alternatives.
Keywords. Banana, genetic improvement, pest management, diseases, abiotic stress factors.
INTRODUCTION
Origin of bananas and plantains



Received: September 26, 2022 / Accepted: October 15, 2022 / Published:15 February 2023
Bananas and plantains belong to the genus Musa and are native to Southeast Asia, including the Indo-Malaysia region and the Australian tropics1 . Therefore, Southeast Asia is the first center of diversity for diploid bananas and plantains2 . Following the introduction of triploid bananas in West Africa and cooking and beer bananas in East Africa, the second center of diversification occurred on this continent, based primarily on mutations3 4 . Thus, the most significant variability of banana cultivars is found in India and Southeast Asia, followed by plantains in Central and West Africa1 .
Botany of bananas and plantains
Bananas and plantains are perennial herbs consisting of an underground stem called a rhizome (commonly called a corm), with roots and vegetative shoots, from which a pseudostem composed of clustered leaf bases protrudes from the ground5 . From the apical meristem, the primordia of the different leaves are distinguished, which grow vertically and differentiate into a leaf base, a petiole, and a blade.
The cultivation cycle varies between 8-12 months, while propagation occurs vegetatively due to the generation of shoots from the corm of the plants. Bananas and plantains are grown in Ecuador, of which many producing countries are parthenocarpy and do not have seeds in the fruit. However, wild species have roots and are essential components in germplasm banks as sources of genes of agricultural importance in conventional and biotechnological breeding programs.
An example is the 'species Calcutta 4' (genotype AA, Musa acuminata ssp. burmaniccoides), recommended for field trials as a highly resistant reference to black sigatoka5 6 7 , used as a parent in conventional breeding programs8 9 , used in molecular studies on plant-pathogen interaction and the discovery of resistance genes to fungal diseases such as black Sigatoka (caused by Pseudocercospora fijiensis10 11 12 13 14 and Panama disease (Fusarium oxysporum f sp. cubense15 16 ).
Inflorescence
At a physiological stage of plant development, after 25 to 50 leaves have been produced, the apical growing point stops producing leaves and becomes an inflorescence. The peduncle grows through the pseudo stem and out the top of the plant. The bracts cover double rows of female nodes with grouped fruits called hands. Each fruit that stands out from each flower is called a finger; later, the bracts will dry up and fall.
The hermaphroditic flowers, usually drooping, are found next to the developing fruit. At the end of the peduncle, the male flowers stand out, full of bracts. This terminal part is called the acorn or male inflorescence5 . Each plant flowers only once, at the end of its life cycle and pollen production. The fertility of cultivated bananas and plantains depends mainly on genetic factors, although environmental factors may also play a role.
Lowering and fruit production, the mother plant dies, which will be replaced by the offspring that develops from the corm, which emerges during the vegetative development of the mother plant.
Origin of edible bananas and plantains
Wild Musa species produce seeds and generally grow wild in forest clearings and along streams. In the past, farmers selected plants with a high level of parthenocarpy (fruit formation without fertilization) because the fruits had fewer seeds due to selection for female sterility, which also helped increase the edibility of this fruit.
These selected plants probably propagated vegetatively by removing suckers from the mother plant. Population movement allowed the dispersal of bananas and plantains to other parts of the world5 . Wild banana species are diploid M. acuminata (AAw) or M. balbisiana (BBw); hybridization between the two wild species occurred in peripheral dry areas of Southeast Asia, where M. balbisiana was endemic when cultivation from M. acuminata started. In addition, triploidy, an essential feature in the evolution of bananas and plantains, was due to the fertilization of a viable diploid ovule formed in interrupted meiosis in the second division with haploid pollen17 .
Most cultivable bananas are triploid (2n= 3X = 33 chromosomes) and are responsible for the world's largest producer of bananas. For example, banana varieties of the Cavendish subgroup (AAA genotypes) are grown in Ecuador, including 'Williams,' 'Gran Enano,' and 'Valery.' On the other hand, there are also diploids (2n = 2X = 22 chromosomes) and tetraploid bananas (2n = 4X = 44 chromosomes), while all wild species of the genus Musa are diploid. Jones (2000) describes a compendium of cultivars divided into genomic groups and ploidy levels5 .
Pollen dispersal and viability
Pollen viability is high in diploid species (88%) and higher in tetraploid species (29%), being nine times lower in cultivable triploids (6%-10%). On the other hand, the triploid cultivar' Gros Michel' (AAA) has 13% viable pollen18 .
Different studies of the cultivar' Valery'19 reported apparent sterility of cultivars of the Cavendish subgroup. However, despite the various difficulties, it has been possible to generate crosses between Cavendish cultivars, obtaining 40 viable embryos from 200 seeds. Then 20,000 pollinated seeds, revealing that the Cavendish cultivars ('Grand Naine' and 'Williams') have low fertility19 .
Sexual and asexual reproduction
Most cultivable bananas and plantains are partially or wholly sterile. Some edible diploids may have female and male sterility due to structural heterozygosity (translocations, inversions), lack of homology between A and B chromosomes, or genetic effects1 . Triploid cultivars have reproductive cells with one or three sets of chromosomes. The plants produce little or no pollen and are partially sterile. They develop fruits without pollination (parthenocarpic); they are seedless and propagate vegetatively. Crossing a triploid (as a female parent) with a diploid (as a male parent) can produce offspring with two, three, or four sets of chromosomes. In those offspring that maintain a uniform set of chromosomes, fertility is re-established; they can produce fruits with seeds when the flowers are pollinated1 . The commercialization of the triploids was possible due to their reduced seed production capacity due to: i) the sterility of the gametes caused by the combination problems generated by the triploid germinative tissue, ii) the irregular or late growth of the pollen tubes in the styles of the female flowers, iii) absence of fertilization even with the development of the tube due to unknown causes; and iv) nectary necrosis at flowering20 .
Conventional breeding programs use the fertility of wild diploid species, but the seeds produced have a low germination rate. At the same time, male infertility and parthenocarpy are closely related, with few exceptions.
Propagation of banana and plantain cultivars is by basal shoots or corms, which develop underground. Each bud is genetically identical to the mother (a clone). Furthermore, although wild banana species are fertile, they propagate vegetatively through shoot production.
Seed dispersal, dormancy, volunteers, invasion, and potential as weeds
Bats perform seed dispersal, as do rodents, squirrels, and monkeys21 1 . In addition, the dormant seeds of some wild bananas are believed to remain viable in the soil for years and can germinate when light passes through the plant bark.
Established populations can be very persistent due to their ability to propagate vegetatively. However, the probability of spontaneously established volunteers is relatively low due to the lack of seeds in cultivars. Edible bananas are not considered invasive and do not compete well with forest species under natural conditions1 . Despite the vegetative reproduction of bananas, suckers may emerge from the remaining corms; however, if insufficient watering and fertilization, the offspring can become stressed and die.
Bananas and plantains are one of the most important crops in the world and are among the top 10 crops for food production. Musaceae are widely distributed in tropical areas and form an essential component in the diet of millions of consumers worldwide. In several countries, bananas and plantains are a staple and leading food; thus, in countries such as Uganda, the average consumption level is 0.5 kg per person per day, rising in some regions to around 1 kg per person per day22 23 .
Bananas and plantain crops represent a sizeable economic export item for Ecuador that reached USD 3,682 million in 2021 (CFN 2022) and, in turn, is a product of high nutritional interest.
These crops grow in humid tropical and subtropical regions, favoring Musaceae to be strongly threatened by pests and diseases. Given rise to the growth of pathogens such as the causal agent of black sigatoka (Pseudocercospora fijiensis), Fusarium (Fusarium oxysporum f. sp. cubense, Foc.), banana bacterial wilt (Xanthomonas campestris pv. musacearum), virus (Banana Bungee Top, Banana Streak Virus), nematodes (Radopholus similis and Pratylenchus coffeae) and insects (Cosmopolites sordidus) which cause significant economic losses worldwide.
All these problems have created a growing demand for new and improved varieties of these crops that respond to the high demand worldwide. This increased consumption highlights the scientific interest in developing and improving this fruit and the constant search for improvements in programs based on yield, quality, and the ability to resist such devastating diseases caused by fungi or other vectors.
There are extensive works published on bananas and plantains, where they deal with issues on the application of genetic engineering to improve characters that confer tolerance to biotic and abiotic stresses and for their biofortification.
The improvement of bananas through conventional crossing techniques continues today to be a process with great difficulties due to the time of the crop cycle between generations, the ploidy levels, the sterility of the edible cultivars, the requirement of physical space, and the limited genetic variation of the species. It is estimated that more than 1000 manual pollinations from 200 plants are needed to obtain around 1000 seeds/year of a selected tetraploid plantain-banana hybrid24 . This estimate makes evident the high degree of labor involved in continuing to depend on traditional techniques for crops such as bananas and plantains without even considering the economic and human cost that is required. It is essential to highlight that, despite these difficulties, it has been possible to obtain hybrids that present genetic resistance to diseases such as black sigatoka and, at the same time, agronomic characteristics of interest.
In 1992, the widespread involvement of important banana plantations by diseases, particularly the fungus Fusarium oxysporum f. sp. cubense called Mal de Panama, and the little research in the search for improved plant material led to the beginning of breeding programs for this crop to obtain resistance to the said pathogen25 . Due to the lack of controls and the threat created by this disease, added to the outbreak of black sigatoka in the early 1970s, genetic techniques began in plant breeding programs to search for varieties with resistance to these diseases. One of the oldest Musa genetic improvement programs still in force is that of the Honduran Foundation for Agricultural Research (FHIA). The United Fruit Company initially established this program for sweet bananas but later included plantains and bananas. Other institutions working actively since the late 1980s and early 1990s on breeding programs are the Center de Coopération Internationale en Recherche Agronomique pour le Développement (CIRAD, France), Center Africain de Recherches Sur Les Bananiers et Plantains (CARBAP, Cameroon), the Internationale Institute of Tropical Agriculture (IITA, Africa), Empresa Brasileira de Pesquisa Agropecuaria (EMBRAPA, Brazil), Center National de Recherches Agronomiques (CNRA, Ivory Coast), the National Center for Research on Banana (NCRB, India), among others3 .
The challenges described above allowed directing the research on Musa spp. towards the field of cell and molecular biology for the improvement of bananas. Plant tissue culture techniques have facilitated germplasm exchange, rapid multiplication, and conservation. Molecular techniques began to be useful for banana breeding due to the use of molecular markers to help in the germplasm management process, select genes and characteristics of interest from a wild population for genetic introgression, and even detect and determine pathogens in banana plants26 .
Traditional banana cultivation and improvement techniques require years of study to evaluate everything from pollination to the generation of the first bunches. These analyses are preliminary tests to determine resistance to pathogens, which may require several crop cycles. Currently, the decreasing costs of molecular assays make these techniques cheaper than the formal study by phenotypes, not to mention the precision and speed when handling many individuals. The development of bioinformatic tools and nucleic acid sequencing technologies for the analysis of genomes of species of agronomic interest is on the rise, facilitating access and understanding of their genetic information, not only to understand and predict the functions of the genes but also to detect regions in the genome that present associations with agronomic characteristics of interest.
Alternative to conventional improvement in Musa
Due to the rapid adaptation of pathogenic organisms to control methods in the field (fungicides, antibiotics, etc.) and the environmental conditions that affect different crops, genetic engineering has become an indispensable tool in various genetic improvements. The ability to modify crops to improve their production, tolerance to environmental changes, and ecological adaptations, among others, are reasons why targeted genetic modification of crops proves to be an essential tool.
Genetic engineering has made it possible to generate genetically modified plants resistant to the burrowing nematode27 to the bacterial wilt of bananas caused by the pathogen Xanthomonas campestris pv. musacearum28 , fusariosis caused by Foc race 429 , among others. Resistance to pathogens and adverse environmental conditions that affect crop development has not been the only approach in which genetic engineering has participated. The biofortification or growth of plants with improved nutritional content has also attracted the attention of researchers. In this area, using a genetic modification of plants has served to obtain a higher provitamin A, known as golden bananas30 Researchers in India and Australia are working on developing banana plants with enriched iron content to alleviate iron deficiency anemia31 . In Ecuador and Belgium, a team of researchers from CIBE-ESPOL and the University of Ghent is developing a biofortified banana plant to increase folate in the fruit (Efrén Santos, personal communication).
Biosafety and the improvement of human health is scientists' primary interest, for which molecular tools have significantly impacted the development of banana plants with resistance to diseases or adaptation to environmental conditions such as drought, high salinity, and high temperatures. In addition, genetic engineering allows us to develop edible vaccines. For this, the banana is an excellent candidate due to the size of its fruit, edible pulp, adequate postharvest handling, great demand worldwide, low production cost, not require cooking for consumption, and proteins maintain their integrity even when cooked at high temperatures, widely planted and harvested in different countries, especially in developing countries. In this sense, in 1992, Charles Arntzen and his colleague Hugh Mason and Colonel Lam of the United States Armed Forces published the first work that began what is now known as edible vaccines. This group of scientists recovered an antigenic molecule (HBsAg, hepatitis B surface antigen) similar to a viral particle from a transgenic tobacco plant32 . This finding could combine genetic engineering and plant molecular biology to create low-cost vaccine production systems. This discovery opened the doors to new research in food crops for synthesizing antigens for various diseases that affect humans and animals. Among these crops, using the fruit of banana plants as a store for edible vaccines seemed to be a great option. However, this crop has problems such as low expression and accumulation of antigens in the fruit, so it must be investigated and optimized33 .
Methods of genetic transformation
The continuous evolution and appearance of new races of pathogens have created the need to accelerate the genetic improvement process to obtain new resistant banana varieties in the short term. To solve the problems that come from having limited genetic variability, low or null fertility, and polyploidy, among others34 , biotechnology has been the tool of choice to deal with this complex situation.
Figure 1. Scheme of the genetic transformation process using Agrobacterium tumefaciens, A) banana cell suspensions (cultivar 'Williams' AAA), B) and C) regeneration of banana embryogenic cells and calluses in ITC-K selection medium after genetic transformation, D) and E) maintenance and cocultivation under controlled light and temperature conditions of genetically transformed banana seedlings, F) GUS histochemical assay on the leaf. (Photo from Santos et. al.)35
In the case of genetic engineering, there are several methods to insert new genes into banana plants to obtain new plants resistant to different types of stress or even to create biofortified fruits36 without altering the characteristics of plant sales. One of the first methodologies used to generate genetically modified banana plants was particle bombardment, a technique that introduces DNA into a living cell by shooting gold or tungsten particles (1-4 µm in diameter), penetrating the cell wall and membranes without being lethal37 . The plant material used are embryogenic calli and embryogenic cell suspensions (ECS) and young tissues to regenerate stably transformed plants. Despite being an efficient method to generate transformed banana plants, this methodology tends to insert multiple copies of DNA into cells, and plants composed of genetically distinct tissues, called chimeras, are also often produced3 .
Using tissues such as embryogenic cells and protoplasts is ideal when avoiding the appearance of possible chimeras. In protoplasts, gene transfer to the cell nucleus is by electroporation, a technique that requires a prior standardization of parameters adjusted to the cultivar with which it will work. The most significant problem in the generation of protoplasts occurs in monocot species 38 .
Another methodology for plant modification is by the soil bacterium Agrobacterium tumefaciens, known to cause tumors in dicotyledonous plants through the transfer of genes located in plasmids present in the Agrobacterium, which are modified to be able to transform plant cells using genes of interest39 . For this same reason, it was not initially used in bananas because, as a monocotyledonous plant, it was not within the host range of the bacterium.
However, May et al. (1995) showed that using a co-culture system of the bacteria and meristematic tissue, they could transform and regenerate genetically modified banana plants, then verify the transformation using the GUS reporter gene and the Southern blot technique40 . A variation of this method is the infiltration of A. tumefaciens employing a vacuum, which was tested by41 in meristematic banana tissue, using the gene encoding neomycin phosphotransferase II (nptII), which generates resistance to the antibiotic kanamycin as a selection tool. Although this variant managed to reduce the mortality rate of the transformed tissue and regenerated a more significant number of plants, it was prone to the generation of chimeras, which it tried to solve (unsuccessfully) by increasing the concentration of antibiotics.
Figure 2. A) In vitro banana plants from the 'Williams' cultivar was used for transformation. B) Meristematic tissue section. C) Cocultivation of the explants in MS medium supplemented with acetosyringone (200 µM) and Agrobacterium tumefacien.s D) Growth of putative transgenic plants two months after the meristematic transformation process.
This Agrobacterium-mediated genetic transformation methodology was standardized and is routinely applied at CIBE-ESPOL to transform Ecuadorian banana and plantain cultivars42 . For which SCE from the cultivar 'Williams' were used in co-culture with the EHA105 strain of A. tumefaciens, the reporter gene uidA was introduced together with the selection gene hpt, which gives resistance to hygromycin (Fig. 3). Green fluorescent protein (GFP) and luciferase (LUC) is also highly used as plant reporter genes.43 44 45 46
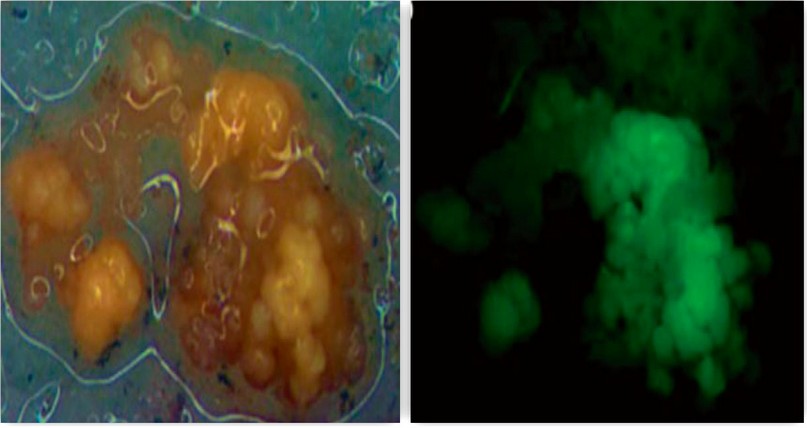
Figure 3. Banana colonies after three months of transformation with the pCAMBIA 1304 and the detection of the GFP reporter gene. (Photo from Santos et., al)47
Commonly, a transgenic organism is associated with environmental biosafety and health risks due to the insertion of foreign DNA. According to Schouten et al. (2006)48 , cisgenesis is very similar to traditional plant breeding because a complete copy of one or more genes of interest, including introns, exons, and promoters, is transferred from sexually compatible organisms. Without altering the recipient species' gene pool and giving them unwanted traits48 .
On the other hand, intragenesis transfers one or several genes. However, these can have genetic elements from different genes and loci, allowing new genetic combinations that can give unique characteristics or desired traits to the modified organism, and which, in turn, like cisgenesis, these genes are from organisms of the same species or family. Due to international regulations on genetically modified organisms that do not discriminate whether an organism is transgenic or cisgenic and the lack of regulatory bodies in this study, future research to improve crop varieties could be delayed48 .
Precision tools for genome editing
The development of reliable and efficient tools that allow making precise changes in the genome of living cells has been a long-awaited desire for researchers involved in life sciences. The controversy generated by using and applying genetically modified crops impacts the favorable development of these technologies for developing cultivars better adapted to biotic and abiotic conditions.
These complications, mainly political and ethical, added to the accelerated growth of biotechnology in recent decades and have allowed the identification and creation of molecular techniques. These new technologies are known as precision genetic engineering technologies, where it is theoretically possible to make changes or edits to the genome of any organism without adding any foreign sequences. Other techniques for editing genetic material incur the problem of being imprecise, and in several cases, it is impossible to determine the insertion site in the genomic sequence.
Additionally, the time it takes to develop new varieties using conventional genetic engineering technologies is considerably high.
Nucleases are a group of enzymes capable of breaking the phosphodiester bond that exists between the monomers of nucleic acids. These enzymes have a standard function during the repair of the genetic material of living beings, having the ability to make breaks in one or both DNA strands. The study of these enzymes has made possible their application in molecular biology for the modification of the genetic material of different organisms. The extensive research and characterization of these enzymatic mechanisms in molecular biology, molecular genetics, and biochemistry have allowed the development of novel tools for their application in genetic improvement and biotechnology, not only in bacteria but also in more complex organisms such as animals and plants. These "new" molecular tools differ from their predecessors, i.e., A. tumefaciens mediated transformation, due to the degree of precision with which they can recognize and manipulate the genetic material of the host organism. These editing tools allow the cutting and joining of DNA molecules at specific sites, creating insertions, deletions, or changes in the host cell's genetic material.
One of the first tools available to scientists was published by Kim et al. (1996), describing the creation of endonucleases capable of recognizing specific sites in the genetic material49 . This group of scientists fused two structural motifs of proteins called zinc fingers to the cutting domain of the endonuclease enzyme Flavobacterium okeanokoites (FokI). The specificity of zinc-finger nucleases (ZFNs) is due to recognizing a set of three nucleotides (triplet) by zinc fingers. Zinc finger proteins (ZFs) of the Cys2-Hys2 type are among eukaryotes' most common DNA-binding motifs and represent the second most encoded protein domain in the human genome50 .
Each Cys2-His2 ZF domain consists of 30 amino acids in a conserved ββα configuration and binds three base pairs of a double-stranded DNA sequence (Liu et al. 1997)51 .
The ability to confer specificity to a protein that binds to DNA at a specific site in a universe of 3.5 million base pairs, as is the case of the human genome, requires at least the recognition of 16-18 base pairs51 . Being able achieved if a protein containing at least 6 ZFs domains is used, and this type of structure was constructed thanks to the discovery of binding sequences, which allow the construction of synthetic ZFs proteins50 .
A ZFNs monomer is composed of 2 distinct functional domains: Cys2-Hys2 ZFs artificial domains at the N-terminus and the FokI non-specific cleavage domain at the C-terminus of the chimeric protein52 . Due to the nature of the FokI endonuclease, it works as a dimer. For this, it needs two binding sites for the ZFs separated by 5-7 bp, which will be the recognition sequence for the cut by the FokI domain.
Each ZFs binding site must be found on one of the targeted DNA strands49 , forming a FokI dimer for cutting both DNA strands. ZFNs have been engineered for gene activation and repression in plants when fused to transcriptional activation or repression domains. Cutting both DNA strands makes it possible to exploit the natural DNA repair mechanisms such as homologous and nonhomologous recombination, allowing the disruption of genes through insertions or deletions typical of repair mechanisms, even achieving the insertion of sequences or processes of complete coding with the help of donor plasmids53 . Since the first report in 1996, ZFNs have been used to manipulate the genome of plants such as Arabidopsis54 , tobacco55 , soybean56 , corn57 , rice58 , and tomato59 . Despite the utility of ZFNs in various crops and organisms, the efficiency of this technology to make specific changes in DNA is usually less than 5%60 .
Another new technology that seeks to identify and localize specific sequences in an organism's genome is transcription activator-like effector-coupled nucleases or TALENs. This precise genome-editing tool works in pairs to search for and cut specific DNA sequences in the same way that ZFNs technology does. Transcriptional activator-like effector proteins recognize and activate specific plant promoters through tandem repeats. In 2007, Römer et al. reported the ability to identify DNA sequences by proteins secreted by the phytopathogenic bacterium Xanthomonas campestris pv61 .
Bosch et al., in 2009, discovered how proteins related to the activation of promoters in host cells could recognize the target sequence in DNA62 . The effector or activator proteins of transcription are found naturally in the genus Xanthomonas bacteria. The domain in charge of recognition is formed by a series of 33-35 amino acids repeated between 15 and 19 times in identical tandems. Each sequence of 33-35 amino acids is responsible for recognizing a nucleotide, where the residues located at positions 12 and 13 of each series confer the specificity of the recognition. This position of high variability is known as the diresidual variable repeat (DVR)52 . The most common diresidual combinations are NI, NG, HD, and NN at the two specific positions mentioned above, which bind to nucleotides A, T, C, and G, respectively 53 . These molecules can change the target organism's DNA when these transcriptional activating effectors fuse with a nuclease domain. The functionality of this technology has been tested in model plants such as Arabidopsis thaliana and Nicotiana benthamiana63 64 65 66 .
One of the most current and novel tools that have quickly become popular in the field of genome editing is the so-called clustered regularly interspaced short palindromic repeats associated with the Cas protein or, in other words, CRISPR/Cas9 (based on the Cas9 protein) for its acronym in English. This new technology has emerged as a faster, cheaper, more accurate, and more efficient option than other precision technologies, such as those mentioned earlier in this chapter.
These palindromic repeats were first found in Escherichia coli by a group of Japanese scientists67 , Although the researchers did not understand their biological role at the time. In 1993, at the University of Alicante, the Spanish researcher Francisco Mojica described palindromic repeats in the halophilic archaea Haloferax mediterranei68 . Mojica himself, together with Janssen in 2002, agreed to coin the term CRISPR to reduce the confusion generated by various acronyms in the scientific community. Several years later, specifically in 2007, Barrangou and colleagues (2007)69 determined that this series of repetitions corresponded to a bacterial immunity system against bacteriophages based on sequences of the pathogen's genetic material. Three types of CRISPR mechanisms and 12 subtypes have been identified so far, which depend on their genetic content structural and content differences70 .
Seen from genetics, cas1 and cas2 are found in all types and subtypes; in Type I, there is cas3; in type II the cas9 gene and in Type III, cas1070 71 . The CRISPR-Cas Type II system is the most studied. This defense system works in such a way that the bacterium integrates into the CRISPR locus of its genome, a small fragment of the infectious virus, called a protospacer, with a length of between 32-38 nucleotides usually. This fragment of the virus's genetic material is incorporated into a new interspaced region after a palindromic repeat at one end of the CRISPR locus and is used by the bacterium to recognize and defend itself from the virus or plasmid in a subsequent attack. The CRISPR locus (the protospacer and part of the CRISPR repeat) is transcribed, and these small RNA fragments generate a CRISPR RNA or crRNA, which is used to guide endonucleases, Cas to the target DNA of the invader based on the complementarity of the spacer sequence72 . The crRNA hybridizes with a second RNA, called a transactivating crRNA (tracrRNA), to form a new complex with the nuclease Cas9. This now mature crRNA, in conjunction with the Cas9 protein, can "seek and recognize" a specific sequence in the host's DNA, thanks to the protospacer sequence, to cut the genetic material of the target organism. The cut in the specified region results from the binding between the crRNA and the protospacer target sequence and a small region called the Protospacer Adjacent Motif (PAM)73 . PAM is a conserved region of between 2-5 nucleotides followed by the 3'-end of the complementary sequence of the crRNA, so important for the nuclease activity of Cas9 that if it is not present, the Cas9:crRNA:tracrRNA complex is not presently capable of recognizing the target sequence.
As described before, the Cas9 enzyme has a molecular scissors-like function capable of cutting both strands of DNA at a specific site in the genome, thanks to the guidance of the crRNA:tracrRNA complex. The Cas9 protein most commonly used for gene editing has been adapted from the Type II CRISPR system of the bacterium Streptococcus pyogenes (Sp)74 . Ongoing studies of the Cas9 enzyme have succeeded in creating three variants for genome editing. The first, the wild-type enzyme Cas9, is capable of cutting double-stranded DNA, resulting in the activation of the cellular repair system for double-stranded damage. There are two mechanisms for repairing these cuts: Nonhomologous DNA End Joining (NHEJ) and Homologous Recombination Repair (HDR)75 . The second variant is mutant versions of Cas9 that have been modified so that they cut a single strand of the target DNA76 . This second variant allows DNA repair by HDR alone because cutting a DNA strand does not activate the NHEJ mechanism. This option allows the design of experiments that use a pair of these mutant complexes to generate two simultaneous cuts at adjacent DNA sites77 . A third variant is called a nuclease deficient Cas9 (dCas9), which has mutations in both the RuvC1 and HNH domains, which inactivate the nuclease activity of the enzyme but do not prevent its ability to bind to target DNA77 78 . This ability to bind uncut DNA has been used to repress or express genes, and if this mutant complex is fused with effector domains, dCas9 can be used to silence or activate gene expression77 71 73 79 80 81 82 .
The gene editing process of the CRISPR-Cas9 system occurs in two steps. The first step is recognizing the specific sequence in the target DNA and cutting the DNA chain, which can be cut in both strands or just one. The second step is DNA repair which follows the action described above. The two repair mechanisms mentioned are the NHEJ and the HDR. The NHEJ repair mechanism causes mutations such as insertions and deletions at the double-strand cleavage site, leading to the silencing of the gene involved. This silencing may be due to a shift in the gene's reading frame or a significant change in the protein-coding region. On the other hand, the HDR repair system can be used to replace a particular sequence at the double-strand break site. This replacement is achieved thanks to homologous recombination guided by a DNA template that performs an insertion, deletion, mutation, correction, or complete replacement of a sequence or gene58 .
The application of the CRISPR-Cas9 gene editing system in plants has been growing in recent years. As mentioned throughout this chapter, the banana is a plant that presents multiple difficulties for its genetic improvement using conventional methodologies. Establishing this molecular tool in bananas allows a step toward applying innovative biotechnological techniques that can be considered in breeding programs. Kaur et al. (2018) presented evidence of phytoene desaturase (PDS) gene editing in the genome of banana cv. 'Rasthali.' They demonstrated the capacity and efficiency of gene editing using the CRISPR-Cas9 system in this plant material, revealing its usefulness for the functional study of genes and use in breeding programs for this crop83 .
Since 2012, the CRISPR-Cas9 system has proven to be a powerful tool that allows the genetic information of various cell types and organisms to be modified. Until now, the versatility and considerable ease of use of the CRISPR-Cas9 system have allowed this tool to be used to modify the genome of organisms such as Streptococcus pneumoniae84 , Escherichia coli84,85 , Lactobacillus reuteri86 , Mycobacterium tuberculosis87 , Saccharomyces cerevisae88 , Aspergillus nidulans89 , Trichoderma reesi90 , Nicotiana benthamiana91 , Arabidopsis thaliana92 , Oryza sativa93 , Solanum lycopersicum94 , Gossypium hirsutum95 , Musa spp.83 , Mus musculus96 , Xenopus laevis97 , Drosophila melanogaster98 , Caenorhabditis elegans99 , among others .
Genetic manipulation applied to banana breeding
Different factors can severely impact banana production since it is a stress-sensitive crop. Banana cultivation can be affected by many biotic factors, such as bacterial, fungal, and viral infections and nematode attacks. Xanthomonas campestris pv. musacearum is a bacterium that threatens production in fields planted with susceptible varieties, specifically in the African continent. Infected plants show rapid yellowing and subsequent wilting of the leaves, affecting the inflorescence and pseudostem, which causes premature fruit ripening100 . This disease is considered of quarantine importance for Latin America, where the best control is not to let the disease enter101 , for which molecular tools have been developed for efficient and specific detection of this bacterium102 . Due to the lack of resistant varieties to this disease, genetic engineering has been considered a strategy for developing resistant varieties103 . Banana plants genetically modified with the ferredoxin-like protein (Pflp) gene from Capsicum annuum, and regulated by the constitutive promoter CaMV35S, have been shown to have up to 100% resistance to the disease compared to infected controls. They develop the typical symptoms of the same due to the existence of a hypersensitivity response, which through programmed cell death (apoptosis), isolates the pathogen, stopping the progress of the infection104 .
The most important fungi in banana are i) Pseudocercospora fijiensis and ii) Fusarium oxysporum f. sp. cubense (Foc). P. fijiensis, the causal agent of black Sigatoka, spread throughout America in the 1980s. This disease produces a progressive yellowing of the leaves and wilting in a few weeks, affecting the fruit's filling and its ripening, causing the cluster to fall105 . In the control of this disease, applications are made with fungicides such as benzimidazoles, triazoles, and others, representing an increase in the selective pressure of the pathogen, increasing resistance to them, leading producers to spray in different cases every 10 to 12 days.
On the other hand, F. oxysporum (with more than 150 unique forms), tropical race 4, represents a severe problem due to its high virulence, almost impossible control, and lack of resistant varieties. This disease is mainly localized in Asia and northern Australia4, recently reported in Colombia and Peru[1]. This pathogen produces yellowing and wilting of the leaves, penetrating through the roots and moving through the xylem until it colonizes the pseudostem, blocking it and causing the death of the plant. There is no effective control of this fungus since it can develop survival structures (chlamydospores) that can remain in the soil for more than 20 years. The best rule is to use resistant/tolerant varieties of the disease. However, there is a shortage of varieties resistant to the new races, and conventional breeding is too slow to deal with it quickly enough.
In both cases, genetic engineering has been considered an effective tool to obtain resistant banana varieties in the short term. Chitinases are a family of proteins that can degrade the chitin present in the cell wall of fungi. Kovács et al. (2013) experimented with overexpression of two chitinases from rice (Oryza sativa) in banana, rcc2, and rcg3, to analyze the resistance of modified plants against P. fijiensis, being the first report of stable transformation in the cultivar' Gros Michel' (AAA)106 . The experiment compared the GM lines produced against a susceptible control,' Gros Michel' (untransformed), and a resistant wild species, 'Calcutta 4' (AA), where most of the GM lines showed a marked average area reduction infected compared to the susceptible variety. A similar approach to finding Foc-resistant varieties is known, where Hu et al. (2013) tested an endo-chitinase gene (chit42) from Trichoderma harzianum in the cultivar 'Furenzhi' (AA) in vitro resistance tests, the genetically modified plants showed a higher level of tolerance compared to non-controls transformed.
Pei and colleagues (2005) transformed banana plants with the gene for a human lysozyme (HL)107 . This enzyme can count on the β-(1,4) glycosidic bond of the peptidoglycan, present in the bacterial cell wall, as well as in the chitin of the fungal cell wall, to obtain plants resistant to FocR4 and bacterial infections. Fifty-one transformed plants were received for greenhouse tests, of which 24 showed tolerance to pathogen infection. Then they were taken to field conditions, where they were infected with the pathogen. Only two of them did not present symptoms, these two being the ones with the highest expression of HL, demonstrating the correlation between the expression of human lysozyme and resistance to FocR4. Currently, there are genetically modified plants of the Cavendish variety that express the genes RGA2 (NB-LRR of Musa acuminata ssp.) and Ced9 (of the nematode Caenorhabditis elegans), which in their majority present resistance to Fusarium oxysporum, even some lines never showed symptoms of Fusarium oxysporum. infection29 .
Genetic engineering for improvement against abiotic stress
The climate change we have experienced in recent years requires obtaining banana varieties that are tolerant to different types of abiotic stress (drought, salinity, low temperatures). Banana is a crop that needs large amounts of water, is susceptible to abiotic factors, and has problems finding varieties resistant to pathogens. For this reason, genetic engineering serves as an instrument to speed up the process of obtaining new types which can cope with possible climate changes and meet food needs.
Shekhawat and collaborators (2011) reported the first study of the generation of transgenic banana plants tolerant to drought and salinity. In this study, they expressed a banana dehydrin (MusaDHN-1), regulating its expression under different types of stress 108 . The transformed events showed an increase in the production of proline in plants (necessary for cellular osmotic adjustment) under abiotic stress conditions, as well as a reduction in the accumulation of malondialdehyde, a marker of oxidative stress, a relative percentage of water in the leaves, which explains in part the correlation of expression of this gene during pressure and the function of dehydrins to protect the lipid membrane. Another gene showing promising results concerning tolerance to abiotic stress is the one belonging to class-10 with pathogenesis (Pathogenesis-Related Class 10; PR10), a group with RNase activity. This gene was isolated from peanut (Arachis hypogaea) and PR10-transformed events also showed increased tolerance to salt stress, with increases in proline production and cell membrane stability compared to control109 .
Aquaporins are a family of proteins that maintain homeostasis and water balance in plant cells. The overexpression of the aquaporin MusaPIP1;2 of the banana cultivar Karibale Monthan', generated an increase in the tolerance to different types of abiotic stress, for they subjected transgenic banana plants to low temperatures (8ºC) for two weeks, drought for another two weeks, and high salinity (250 mM NaCl) for ten days. The physiological parameters for all the tests were photosynthetic capacity, relative water content, and proline production, among others, to measure the behavior and response of all events against stress factors. In all the analyses, the genetically modified plants showed a higher tolerance than the non-transformed control, demonstrating the relationship of these aquaporins with the ability of plants to respond favorably under stress conditions. There are similar experiments overexpressing the aquaporin Musa COP26, which improves the tolerance of transgenic bananas to salinity stress110,111 .
The genetic transformation from 'Williams' banana embryogenic suspensions at CIBE used genes such as ADP 1 (SaARF1) from Spartina alterniflora, a grass that grows in coastal areas and is known to prevent soil erosion in areas with high salinity112 . This gene has proven effective in rice cultivation, conferring tolerance to saline and drought stress to the events that express it113 . The regenerated plants are currently in the in vitro multiplication phase to perform the respective tests (Fig. 4).
Figure 4. Banana plants (cultivar 'Williams', AAA) genetically modified by Agrobacterium tumefaciens in the in vitro multiplication phase.
Genetic engineering for biofortification
Finally, a marked trend in recent years is the biofortification of crops, and bananas are no exception, given the need in countries where micronutrient deficiencies result from unbalanced diets114 . Plantains from the Cavendish group are genetically modified using the MtPsy2a and ZmPsy1 genes (used in Golden rice) to produce a minimum β-carotenoid concentration of 20 µg/g dry matter, rising to 55 µg/g. g in one of the transgenic events expressing MtPsy2a. However, plants that overexpressed ZmPsyI showed undesirable phenotypes, such as the presence of "golden" young leaves (they returned to their greenish color over time), "golden" clusters (with the ripening of the fruit, the color changed to a mixture between green and orange) and fruit pulp with a coloration that went from intense yellow to orange.
The production of β-carotenoids proved stable in different generations of transgenic lines30 . Human studies are underway with the "golden" banana in the United States of America, a project funded by the Bill and Melinda Gates Foundation 36 .
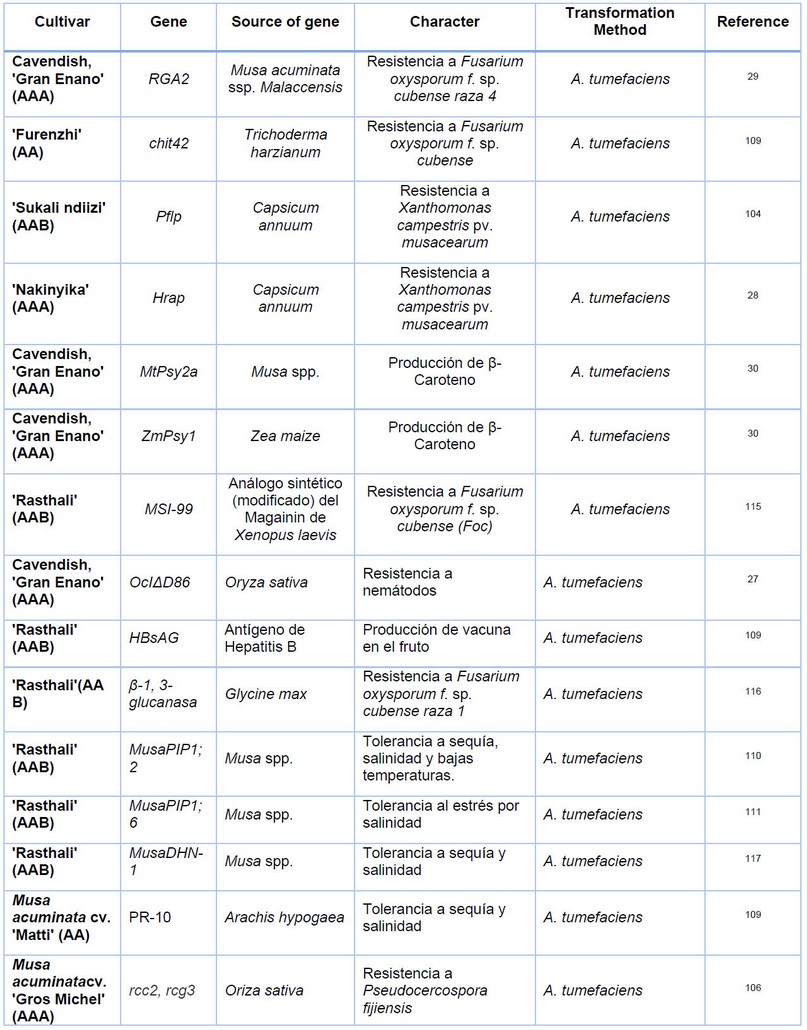
Table 1. Banana and plantain cultivars are improved by genetic transformation.
Recently, at CIBE-ESPOL, a research project is being carried out using the GTPCHI genes of bananas and the ADCS of Arabidopsis thaliana in collaboration with the Laboratory of Functional Biology in Plants of the University of Ghent, led by Professor Dominique Van Der Straeten. The genes were fused with the promoter of the expansin gene in bananas, active in fruit tissues (Efrén Santos personal communication). Table 1 summarizes different works published on genetic engineering in Musa species.
CONCLUSION
Genetic engineering is a tool used worldwide for the improvement of crops. In species of the Musa genus, there are many studies carried out for genetic modification for disease resistance and biofortification. Recently, genome editing has been used massively in crop improvement, and bananas are no exception. Although there are few studies published to date, it is expected to increase in the coming years. Among the pending challenges is allowing the cultivation of genetically modified bananas and plantains for food security and commercial purposes.
REFERENCES
1. Nuez, F. M.S. Andersson and MC de Vicente: Gene flow between crops and their wild relatives. Genetic Resources and Crop Evolution 2011 58:4 58, 619–619 (2011).
2. Daniells, J. W., Jenny, C., Karamura, D. A. & Tomekpe, K. Musalogue: a catalogue of Musa germplasm. Diversity in the genus Musa. Cirad 213 (2001) doi:10.3/JQUERY-UI.JS.
3. Pillay, M., Tenkouano, A., Ude, G. & Ortiz, R. Molecular characterization of genomes in Musa and its applications. Banana improvement: cellular, molecular biology, and induced mutations. Proceedings of a meeting held in Leuven, Belgium, 24-28 September 2001 271–286 (2004).
4. Nelson, S., Ploetz, R. & Kepler, A. Musa species (banana and plantain). Species profiles for pacific island agro forestry 2006, (2006).
5. Muirhead, I.F. y Jones, D. R. Diseases of Banana, Abacá and Enset. Postharvest disease 50, 190–206 (2000).
6. Orjeda, G. Evaluating bananas: a global partnership. Results of IMTP Phase II. International Network for the Improvement of Banana and Plantain, Montpellier, France. ISBN.
7. Carlier, J. (Jean) & International Network for Improvement of Banana and Plantain. Global evaluation of Musa germplasm for resistance to Fusarium wilt, Mycosphaerella leaf spot diseases, and nematodes : in-dept evaluation. 63 (2002).
8. Rosales, F. E. & Pocasangre, L. E. Mejoramiento convencional de banano y plátano: estrategias y logros [Conventional banana and plantain breeding: strategies and achievements]. (2002).
9. Menon, R. Banana breeding. Banana: Genomics and Transgenic Approaches for Genetic Improvement 13–34 (2016) doi:10.1007/978-981-10-1585-4_2.
10. Zandjanakou-Tachin, M. et al. Identification and genetic diversity of Mycosphaerella species on banana and plantain in Nigeria. Plant Pathol 58, 536–546 (2009).
11. Azhar, M. & Heslop-Harrison, J. S. Genomes, diversity and resistance gene analogues in Musa species. Cytogenet Genome Res 121, 59–66 (2008).
12. Miller, R. N. G. et al. Analysis of non-TIR NBS-LRR resistance gene analogs in Musa acuminata Colla: Isolation, RFLP marker development, and physical mapping. BMC Plant Biol 8, 15 (2008).
13. N Passos, M. A. et al. Development of expressed sequence tag and expressed sequence tag–simple sequence repeat marker resources for Musa acuminata. AoB Plants 2012, 30 (2012).
14. Sánchez Timm, E. et al. Identification of Differentially-Expressed Genes in Response to Mycosphaerella fijiensis in the Resistant Musa Accession ‘Calcutta-4’ Using Suppression Subtractive Hybridization. PLoS One 11, (2016).
15. Swarupa, V., Ravishankar, K. v. & Rekha, A. Plant defense response against Fusarium oxysporum and strategies to develop tolerant genotypes in banana. Planta 239, 735–751 (2014).
16. Ravishankar, K. v. et al. Genetic diversity and population structure analysis of mango (Mangifera indica) cultivars assessed by microsatellite markers. Trees - Structure and Function 29, 775–783 (2015).
17. Simmonds, N.W. 1962. The evolution of the bananas. Tropical Science Series. Longmans, London (GBR). 170p.
18. Fortescue, J. A. & Turner, D. W. Pollen fertility in Musa: Viability in cultivars grown in Southern Australia. Aust J Agric Res 55, 1085–1091 (2004).
19. Aguilar Morán, J. F. Improvement of cavendish banana cultivars through conventional breeding. Acta Hortic 986, 205–208 (2013).
20. Citogenética y mejoramiento genético del banano (Musa spp.). - Portal Embrapa. https://www.embrapa.br/busca-de-publicacoes/-/publicacao/641469/citogenetica-e-melhoramento-genetico-da-bananeira-musa-spp.
21. Meng, L., Gao, X., Chen, J. & Martin, K. Spatial and temporal effects on seed dispersal and seed predation of Musa acuminata in southern Yunnan, China. Integr Zool 7, 30–40 (2012).
22. Komarek, A. The determinants of banana market commercialization in Western Uganda. Afr J Agric Res 5, 775–784 (2010).
23. Smale, M. et al. An Economic Assessment of Banana Genetic Improvement and Innovation in the Lake Victoria Region of Uganda and Tanzania. doi:10.2499/9780896291645RR155.
24. Ortiz, R, Vuylsteke D (1993) The genetics of black sigatoka resistance, growth and yield parameters in 4x and 2x plantain-banana hybrids En: Ganry J (ed.) breeding banana and plantain for resistance to diseases and pests, p. 379. CIRAD- INIBAP. Montpellier.
25. Kumar, J., Chaube, H., Singh, U. & Mukhopadhyay, A. Plant diseases of international importance. Volume III. Diseases of fruit crops. Undefined (1992).
26. Ortiz, R. (2013). Conventional banana and plantain breeding. Acta Hortic. 986, 177-194 DOI:10.17660/ActaHortic.2013.986.19.
27. Atkinson, H. J., Grimwood, S., Johnston, K. & Green, J. Prototype demonstration of transgenic resistance to the nematode Radopholus similis conferred on a banana by a cystatin. Transgenic Res 13, 135–142 (2004).
28. Tripathi, L., Mwaka, H., Tripathi, J. N. & Tushemereirwe, W. K. Expression of sweet pepper Hrap gene in banana enhances resistance to Xanthomonas campestris pv. musacearum. Mol Plant Pathol 11, 721–731 (2010).
29. Dale, J. et al. Transgenic Cavendish bananas with resistance to Fusarium wilt tropical race 4. Nat Commun (2017) doi:10.1038/s41467-017-01670-6.
30. Paul, J. Y. et al. Golden bananas in the field: elevated fruit pro-vitamin A from the expression of a single banana transgene. Plant Biotechnol J 15, 520–532 (2017).
31. Patel, P., Yadav, K. & Ganapathi, T. R. Biofortification for alleviating iron deficiency anemia. Banana: Genomics and Transgenic Approaches for Genetic Improvement 301–337 (2016) doi:10.1007/978-981-10-1585-4_20.
32. McQueen-Mason, S., Durachko, D. M. & Cosgrove, D. J. Two Endogenous Proteins That Induce Cell Wall Extension in Plants. Plant Cell (1992) doi:10.2307/3869513.
33. Arntzen, C. Plant‐made pharmaceuticals: from 'Edible Vaccines' to Ebola therapeutics. Plant Biotechnol J 13, 1013 (2015).
34. Genetic Transformation of Perennial Tropical Fruits on JSTOR. https://www.jstor.org/stable/4293771.
35. Santos, E. et al. Advances in banana transformation through Agrobacterium tumefaciens in Ecuador: Progress, challenges, and perspectives. in Acta Horticulturae vol. 1114 197–202 (International Society for Horticultural Science, 2016).
36. Waltz, E. Vitamin A Super Banana in human trials. Nat Biotechnol 32, 857 (2014). https://doi.org/10.1038/nbt0914-857.
37. Sanford, J. C., Klein, T. M., Wolf, E. D. & Allen, N. Delivery of substances into cells and tissues using a particle bombardment process. http://dx.doi.org/10.1080/02726358708904533 5, 27–37 (2007).
38. Roderick, H., Tripathi, L. & Poovarasan, S. Transgenic Approaches to Improve Resistance to Nematodes and Weevils. Undefined 247–260 (2016) doi:10.1007/978-981-10-1585-4_17.
39. Hoekema, A., Hirsch, P. R., Hooykaas, P. J. J. & Schilperoort, R. A. A binary plant vector strategy based on separation of vir- and T-region of the Agrobacterium tumefaciens Ti-plasmid. Nature vol. 303 179–180 Preprint at https://doi.org/10.1038/303179a0 (1983).
40. Hard, T., Barnes, H., Larsson, C., Gustafsson., Lund, J. © 199 5 Nature Publishing Group http://www.nature.com/nsmb. Nature 2, 983–989 (1995).
41. Acereto-Escoffié, P. O. M. et al. Agrobacterium-mediated transformation of Musa acuminata cv. 'Grand Nain' scalps by vacuum infiltration. Sci Hortic 105, (2005).
42. Luis Eduardo Sánchez & Ordoñez, E. G. S. Estandarización del protocolo de transformación genética de células embriogénicas de banano de la variedad ‘Williams’ (AAA) mediada por Agrobacterium tumefaciens. Revista Tecnológica - ESPOL 23, (2010).
43. Santos, E. et al. Tagging novel promoters in banana using the luciferase reporter gene. in Acta Horticulturae vol. 763 99–105 (2007).
44. Santos Ordóñez EG. Characterization and isolation of T-DNA tagged banana promoters active during in vitro regeneration and low temperature stress. (Katholieke Universiteit Leuven, 2008).
45. Santos, E. et al. Characterization and isolation of a T-DNA tagged banana promoter active during in vitro culture and low temperature stress. BMC Plant Biol 9, 77 (2009).
46. Villao, L. et al. Activity characterization of the plantain promoter from the heavy metal-associated isoprenylated plant gene (MabHIPP) using the luciferase reporter gene. Plant Gene 19, (2019).
47. Santos, E. et al. Promoter Analysis in Banana BT - Banana: Genomics and Transgenic Approaches for Genetic Improvement. in (eds. Mohandas, S. & Ravishankar, K. v) 157–179 (Springer Singapore, 2016). doi:10.1007/978-981-10-1585-4_11.
48. Schouten, H. J., Krens, F. A. & Jacobsen, E. Cisgenic plants are similar to traditionally bred plants: International regulations for genetically modified organisms should be altered to exempt cisgenesis. EMBO Rep 7, 750–753 (2006).
49. Kim, Y. G., Cha, J. & Chandrasegaran, S. Hybrid restriction enzymes: Zinc finger fusions to Fok I cleavage domain. Proc Natl Acad Sci U S A 93, (1996).
50. Gaj, T., Gersbach, C. A. & Barbas Iii, C. F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol 31, 397–405 (2013).
51. Liu, Q., Segal, D. J., Ghiara, J. B. & Barbas, C. F. Design of polydactyl zinc-finger proteins for unique addressing within complex genomes. Proc Natl Acad Sci U S A 94, (1997).
52. Kamburova, V. S. et al. Genome Editing in Plants: An Overview of Tools and Applications. International Journal of Agronomy vol. 2017 Preprint at https://doi.org/10.1155/2017/7315351 (2017).
53. Liu, W. & Stewart, C. N. Plant synthetic promoters and transcription factors. Curr Opin Biotechnol 37, 36–44 (2016).
54. Zhang, F. et al. High frequency targeted mutagenesis in Arabidopsis thaliana using zinc finger nucleases. Proc Natl Acad Sci U S A 107, (2010).
55. Townsend, J. A. et al. High-frequency modification of plant genes using engineered zinc-finger nucleases. Nature 459, (2009).
56. Curtin, S. J. et al. Targeted mutagenesis of duplicated genes in soybean with zinc-finger nucleases. Plant Physiol 156, (2011).
57. Shukla, V. K. et al. Precise genome modification in the crop species Zea mays using zinc-finger nucleases. Nature 459, (2009).
58. Zhang, Y. et al. An A20/AN1-type zinc finger protein modulates gibberellins and abscisic acid contents and increases sensitivity to abiotic stress in rice (Oryza sativa). J Exp Bot 67, (2016).
59. Hilioti, Z., Ganopoulos, I., Ajith, S., Bossis, I. & Tsaftaris, A. A novel arrangement of zinc finger nuclease system for in vivo targeted genome engineering: the tomato LEC1-LIKE4 gene case. Plant Cell Rep 35, (2016).
60. Ainley, W. M. et al. Trait stacking via targeted genome editing. Plant Biotechnol J 11, (2013).
61. Römer, P. et al. Plant pathogen recognition mediated by promoter activation of the pepper Bs3 resistance gene. Science (1979) 318, (2007).
62. Boch, J. et al. Breaking the code of DNA binding specificity of TAL-type III effectors. Science (1979) 326, (2009).
63. Zhang, Y. et al. Transcription activator-like effector nucleases enable efficient plant genome engineering. Plant Physiol 161, (2013).
64. Mahfouz, M. M. & Li, L. TALE nucleases and next generation GM crops. GM Crops 2, (2011).
65. Christian, M., Qi, Y., Zhang, Y. & Voytas, D. F. Targeted Mutagenesis of Arabidopsis thaliana Using Engineered TAL Effector Nucleases. G3: Genes, Genomes, Genetics 3, (2013).
66. Cermak, T. et al. Erratum: Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting (Nucleic Acids Research (2011) 39 (e82) DOI: 10.1093/nar/gkr218). Nucleic Acids Research vol. 39 Preprint at https://doi.org/10.1093/nar/gkr739 (2011).
67. Ishino, Y., Shinagawa, H., Makino, K., Amemura, M. & Nakatura, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isoenzyme conversion in Escherichia coli, and identification of the gene product. J Bacteriol 169, (1987).
68. Mojica, F. J. M., Juez, G. & Rodriguez‐Valera, F. Transcription at different salinities of Haloferax mediterranei sequences adjacent to partially modified PstI sites. Mol Microbiol 9, (1993).
69. Barrangou, R. et al. CRISPR provides acquired resistance against viruses in prokaryotes. Science (1979) 315, (2007).
70. Barrangou, R. & Marraffini, L. A. CRISPR-cas systems: Prokaryotes upgrade to adaptive immunity. Molecular Cell vol. 54 Preprint at https://doi.org/10.1016/j.molcel.2014.03.011 (2014).
71. Fichtner, F., Urrea Castellanos, R. & Ülker, B. Precision genetic modifications: A new era in molecular biology and crop improvement. Planta vol. 239 Preprint at https://doi.org/10.1007/s00425-014-2029-y (2014).
72. Brouns, S. J. J. et al. Small CRISPR RNAs guide antiviral defense in prokaryotes. Science (1979) 321, (2008).
73. Jinek, M. et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science (1979) 337, 816–821 (2012).
74. Hsu, P. D. et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat Biotechnol 31, (2013).
75. Sander, J. D. & Joung, J. K. CRISPR-Cas systems for editing, regulating and targeting genomes. Nature Biotechnology vol. 32 Preprint at https://doi.org/10.1038/nbt.2842 (2014).
76. Nishimasu, H. et al. Crystal structure of Cas9 in complex with guide RNA and target DNA. Cell 156, (2014).
77. Chen, X. et al. In trans paired nicking triggers seamless genome editing without double-stranded DNA cutting. Nat Commun 8, (2017).
78. Gasiunas, G., Barrangou, R., Horvath, P. & Siksnys, V. Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc Natl Acad Sci U S A 109, (2012).
79. Gilbert Luke A et al. CRISPR-mediated modular RNA-guided regulation of transcription in eukaryotes. Cell 154, (2013).
80. Lowder, L. G., Malzahn, A. & Qi, Y. Plant gene regulation using multiplex CRISPR-dCas9 artificial transcription factors. in Methods in Molecular Biology vol. 1676 (2018).
81. Perez-Pinera, P. et al. RNA-guided gene activation by CRISPR-Cas9-based transcription factors. Nat Methods 10, (2013).
82. LS, Q. et al. Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression. Cell 184, (2021).
83. Kaur, N. et al. CRISPR/Cas9-mediated efficient editing in phytoene desaturase (PDS) demonstrates precise manipulation in banana cv. Rasthali genome. Funct Integr Genomics 18, (2018).
84. Jiang, W., Bikard, D., Cox, D., Zhang, F. & Marraffini, L. A. RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nat Biotechnol 31, (2013).
85. Cong, L. et al. Multiplex genome engineering using CRISPR/Cas systems. Science (1979) 339, (2013).
86. Oh, J. H. & van Pijkeren, J. P. CRISPR-Cas9-assisted recombineering in Lactobacillus reuteri. Nucleic Acids Res 42, (2014).
87. Choudhary, E., Thakur, P., Pareek, M. & Agarwal, N. Gene silencing by CRISPR interference in mycobacteria. Nat Commun 6, (2015).
88. Ronda, C. et al. CrEdit: CRISPR mediated multi-loci gene integration in Saccharomyces cerevisiae. Microb Cell Fact 14, (2015).
89. Nødvig, C. S., Nielsen, J. B., Kogle, M. E. & Mortensen, U. H. A CRISPR-Cas9 system for genetic engineering of filamentous fungi. PLoS One 10, (2015).
90. Liu, R., Chen, L., Jiang, Y., Zhou, Z. & Zou, G. Efficient genome editing in filamentous fungus Trichoderma reesei using the CRISPR/Cas9 system. Cell Discov 1, (2015).
91. Nekrasov, V., Staskawicz, B., Weigel, D., Jones, J. D. G. & Kamoun, S. Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nature Biotechnology vol. 31 Preprint at https://doi.org/10.1038/nbt.2655 (2013).
92. Li, J. F. et al. Multiplex and homologous recombination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nature Biotechnology vol. 31 Preprint at https://doi.org/10.1038/nbt.2654 (2013).
93. Belhaj, K., Chaparro-Garcia, A., Kamoun, S. & Nekrasov, V. Plant genome editing made easy: Targeted mutagenesis in model and crop plants using the CRISPR/Cas system. Plant Methods vol. 9 Preprint at https://doi.org/10.1186/1746-4811-9-39 (2013).
94. Shimatani, Z. et al. Targeted base editing in rice and tomato using a CRISPR-Cas9 cytidine deaminase fusion. Nat Biotechnol 35, (2017).
95. Li, C., Unver, T. & Zhang, B. A high-efficiency CRISPR/Cas9 system for targeted mutagenesis in Cotton (Gossypium hirsutum L.). Sci Rep 7, (2017).
96. Hirose, M. et al. CRISPR/Cas9-mediated genome editing in wild-derived mice: Generation of tamed wild-derived strains by mutation of the a (nonagouti) gene. Sci Rep 7, (2017).
97. Wang, F. et al. Targeted gene disruption in Xenopus laevis using CRISPR/Cas9. Cell and Bioscience vol. 5 Preprint at https://doi.org/10.1186/s13578-015-0006-1 (2015).
98. Bassett, A. R., Tibbit, C., Ponting, C. P. & Liu, J. L. Highly Efficient Targeted Mutagenesis of Drosophila with the CRISPR/Cas9 System. Cell Rep 4, (2013).
99. Waaijers, S. et al. CRISPR/Cas9-targeted mutagenesis in Caenorhabditis elegans. Genetics vol. 195 Preprint at https://doi.org/10.1534/genetics.113.156299 (2013).
100. Tripathi, L. et al. Xanthomonas wilt: A threat to banana production in East and Central Africa. Plant Dis 93, (2009).
101. Manzo-sánchez, G., Ciencias, F. de & Colima, U. de. Diseases of quarantine and economic importance in banana tree (Musa sp.) in México. Revista Mexicana de Fitopatología 32, 89–107 (2014).
102. Adikini, S. et al. Development of a specific molecular tool for detecting Xanthomonas campestris pv. musacearum. Plant Pathol 60, (2011).
103. Tripathi, L., Tripathi, J. N. & Tushemereirwe, W. K. Strategies for resistance to bacterial wilt disease of bananas through genetic engineering. African Journal of Biotechnology vol. 3 Preprint at (2004).
104. Namukwaya, B. et al. Transgenic banana expressing Pflp gene confers enhanced resistance to Xanthomonas wilt disease. Transgenic Res 21, (2012).
105. F., G. [Epidemiology and ecology of the Sigatoka negra (Mycosphaerella fijiensis, Morelet) in bananas (Musa sp.), in Costa Rica]. [Spanish]. (1990) doi:10.3/JQUERY-UI.JS.
106. Kovács, G. et al. Expression of a rice chitinase gene in transgenic banana ('Gros Michel', AAA genome group) confers resistance to black leaf streak disease. Transgenic Res 22, 117–130 (2013).
107. Pei, X. W. et al. Creation of transgenic bananas expressing human lysozyme gene for Panama wilt resistance. J Integr Plant Biol 47, (2005).
108. Shekhawat, U. K. S., Srinivas, L. & Ganapathi, T. R. MusaDHN-1, a novel multiple stress-inducible SK(3)-type dehydrin gene, contributes affirmatively to drought- and salt-stress tolerance in banana. Planta 234, 915–932 (2011).
109. Rustagi, A. et al. High Efficiency Transformation of Banana [Musa acuminata L. cv. Matti (AA)] for Enhanced Tolerance to Salt and Drought Stress Through Overexpression of a Peanut Salinity-Induced Pathogenesis-Related Class 10 Protein. Mol Biotechnol (2014) doi:10.1007/s12033-014-9798-1.
110. Sreedharan, S., Shekhawat, U. K. S. & Ganapathi, T. R. Transgenic banana plants overexpressing a native plasma membrane aquaporin MusaPIP1;2 display high tolerance levels to different abiotic stresses. Plant Biotechnol J 11, 942–952 (2013).
111. Sreedharan, S., Shekhawat, U. K. S. & Ganapathi, T. R. Constitutive and stress-inducible overexpression of a native aquaporin gene (MusaPIP2;6) in transgenic banana plants signals its pivotal role in salt tolerance. Plant Mol Biol 88, 41–52 (2015).
112. Joshi, R. et al. Salt adaptation mechanisms of halophytes: Improvement of salt tolerance in crop plants. in Elucidation of Abiotic Stress Signaling in Plants: Functional Genomics Perspectives, Volume 2 (2015). doi:10.1007/978-1-4939-2540-7_9.
113. Joshi, R., Ramanarao, M. V., Lee, S., Kato, N. & Baisakh, N. Ectopic expression of ADP ribosylation factor 1 (SaARF1) from smooth cordgrass (Spartina alterniflora Loisel) confers drought and salt tolerance in transgenic rice and Arabidopsis. Plant Cell Tissue Organ Cult 117, (2014).
114. de Valença, A. W., Bake, A., Brouwer, I. D. & Giller, K. E. Agronomic biofortification of crops to fight hidden hunger in sub-Saharan Africa. Global Food Security vol. 12 Preprint at https://doi.org/10.1016/j.gfs.2016.12.001 (2017).
115. Chakrabarti, A., Ganapathi, T. R., Mukherjee, P. K. & Bapat, V. A. MSI-99, a magainin analogue, imparts enhanced disease resistance in transgenic tobacco and banana. Planta 216, (2003).
116. Maziah, M., Sariah, M. & Sreeramanan, S. Transgenic banana Rastali (AAB) with β-1, 3-glucanase gene for tolerance to fusarium wilt race 1 disease via Agrobacterium-mediated transformation system. Plant Pathol J (Faisalabad) 6, (2007).
117. Shekhawat, U. K. S., Srinivas, L. & Ganapathi, T. R. MusaDHN-1, a novel multiple stress-inducible SK(3)-type dehydrin gene, contributes affirmatively to drought- and salt-stress tolerance in banana. Planta 234, 915–932 (201
Citation: Villao, L. , Chávez, T. , Pacheco, R. , Sánchez, E. , Bonilla J. , Santos, E. Genetic improvement in Musa through modern biotechnological methods. Revis Bionatura 2023;8 (1)20. http://dx.doi.org/10.21931/RB/2023.08.01.20