2020.05.01.5
Files > Volume 5 > Vol 5 No 1 2020
INVESTIGATION / RESEARCH
Morphological study of different varieties of rice traits influencing nitrogen and water uptake efficiency

Raghad S. Mouhamad
Available from: http://dx.doi.org/10.21931/RB/2020.05.01.5
ABSTRACT
This research aimed at establishing the morphology of the root and sizes under various irrigation cultures. The comparison was made for root to shoot ratio under the traditional culture of flooding. We hypothesize that, due to limited root system development under aerobic conditions, rice is poorly adapted to different environments. In the meantime, there has to be an increase in demand for grain and output per area, as newly planted areas are scarce. This study discusses the latest theoretical physiological, metabolic and genetic factors affecting nitrogen intake and use in different N processes. It covers the root system's position and nitrate- ammonium assimilation and its relationship with Nitrogen Use Efficiency (NUE) and Water Use Efficiency (WUE), were discussed. Phenotyping and molecular breeding techniques concerning N and water regimes and genetic diversity were also evaluated and simulated.
Keywords: QTLs, NUE, WUE, Morphology, Irrigation culture
INTRODUCTION
Rice root is a fasciculate system and sense at the herbaceous plant, chill, and water deficiencies sensitive 1; 2, indices, the essential role of the root is extracted and absorption of dissolved minerals and water from the flooded region 3; 4. Recently studies report about the characteristics of root for assessed and attributed the relationships genetic expression and root physiology and morphology 5; 6. The reactions of nitrogen (N) to organic growth and associated characteristics can, therefore, reflect the genetic traits inherited from ancestral to cultivated species 7; 8; 46. The features to improve root for uptake N catch associated with profundity, density 9; 10. The regular plant irrigation, including alternative wetting and mild soil drying drainage, could improve root tip cell ultra-structure; enhance root thickness size and cytokinin accumulation in root 11. Root architecture traits have been related to water and N uptake such as the ammonium (NH4+) transport systems at physiological levels, exhibit linear kinetics 12; 13. Ammonium: nitrate (NH4+: NO3-) ratio arrived at 1:100 in agriculture land, with low focus NH4+ uptake from plant roots highest rates (14). Unusually, perhaps the root plasma film negative electrical polarization in this way its high fondness to the NH4+ more than NO3- 15; 16. Without a doubt, rice root has NH4+ tolerant respiratory increment, and show neither a tight electrochemical angle for NH4+ efflux over the plasma layer 17. The efflux components of transport NH4 +, because of the intense cytosolic concentration of NH4 + found under NH4 + conditions, are often energy-consuming due to inward plant plasma membrane. 18. Because of the enormous size of NH4 +, the energy consumption in respiratory oxygen could increase 19; 20. But deep root systems extract more water from dee phrenic soil layers as a result of changes in as similar partitioning, where roots grow more (number and depth) during vegetative growth. Partitioning between different shooting components is seldom hindered 21. aerobic rice farming is a disruptive technology that aims to reduce water consumption, but the vulnerability of rice to aerobic conditions has restricted its progress 22. Total root biomass variability was mainly due to the individual root growth for aerobic culture. In aerobics, the stomach closing was distinct at the vegetative stage although the soil water production was close to the field power, in part due to poor rooting, vigor 23. Nitrogen fertilizer as an Ammonium or nitrate is associated with a release of Green House Gases (GHGs) arrived at 10- 20% from N fertilizer uses in the world 24. Alternative agronomic approaches can be established for minimizing the use of N-fertilizers through the production and adoption of NUE varieties 25, the use of molecular breeding inputs, agronomy and nutrient modeling, genetic variation 26. The usability of NO3-N seems, therefore, to be impaired in the NUE as well, representing the condition of nature, two carriers family nitrate 1/peptide carrier family (NPF) and 2 families of nitrate carriers (NRT2) were response on uptake and translocation of nitrate in rice 27. It is still NUE is only about 50-fold 28, which indicates that there is a big challenge to develop rice plants with a strong NUE capacity 29. For understanding the pathway regular of N movement in rice root and increasing NUE results from Quality of N-use (NUtE) & quality of N-uptake (NUpE) by increase lateral roots to absorbed more surface area for N uptake 30. Script enzymes lead to physiology and quantitative genetics in hereditary strategy enhanced proteins that connect N translocations of quantitative traits loci (QTLs) 31. Obara et al. 32 found in the study. Several QTLs were mapped to chromosome regions containing GS2 in rice for agronomic features related to the use and yield of N.
Water management had the association between the root morphological characteristics and the productivity of water use in rained lowlands, deep water, marshland, and rain-fed uplands and irrigated grain 33. They review the findings of many studies of SRI supporters. It is more important for poor and marginalized farmers because it tends to boost yields in terms of increased radically-based physiological productivity 34. Nature rice plants become likely to develop a variety of climatic conditions, including sustained aerobic, drought (radical or irregular) 47, poor soil quality, and floods 35. Hence, the classification among root epigenetic regulation characteristics desirable for versatility to the unique number of conditions encountered by rice grains, and even the hereditary territories accountable for such plasticity characteristics, can encourage the choice for large tolerance of rice genotypes to variable conditions to maintain reliable output 36.
MATERIAL AND METHODS
Case 1#
Simulated results of the study by Matsunami et al. 36 for four Indica rice varieties (Puluik Arang, Badari Dhan, Shwe Nang Gyi, and Ratul), after transplanting varieties were identified under flooded condition (soil water potentials of -0.02 Megapascals (MPa)) 43% [w/w], and unflooded soil type (probable soil water; -0.10 MPa), 33% [w/w]. Water consumption became assessed about three times every day. Therefore, shooting and root biomass were evaluated at three weeks by extracting all the root and shoot and drying at 80 ° C for more than three days and weighing. Root statistical analysis: measurement of the firing ratio in SAS software (version 6.12, SAS Institute, Cary, NC, USA) and graphed for each case at the excel system 2010.
Case 2#
This simulated Qun et al. 41 survey analyzing two elite check rice (CK) cultivars (SY-63 and HD-5) and super rice (IIY-084 and WYJ-24) varieties and water quality. The two cultivars and varieties mature in pots with three soil moisture levels after 11 days from transplantation to maturity, three water specifications have been placed by regulating water distribution in well-watered (WW) (0) kilopascal (kPa), mild water deficit (MWD) (–15±5) kPa and severe water deficit (SWD) (–30±5) kPa. Four pots of each the root and shoot dry weight (gpot-1) measurement has been dried at 70 ° C to continue in the oven during drought. Root: shoot ratio was calculated using the formulas Root/Shoot and Painted on the adoption of the program of Excel 2010.
Case 3#
Chu et al. 44 Two representative cultivars (Chunyou927 (CY-927) and Yongyou538 (YYY-537)) and two representative JIR cultivars (Xiushui09 (XS-09)) and Zhejing99(ZJ-99)) have been searched for in the field of alternative wetters and severe drying experiments (AWSD), Watered up to 30 kPa of surface capacity and constant flooding with 2-3 cm of precipitation, CF. Precipitation is continually flooded. Upon physiological maturity, the dry matter of each root and shoot was estimated at a constant weight upon drying at 70 ° C and then weighed and measured by Root: shooting ratio equations for IJHR cultivars in addition to the root of the IJHR: efficiency of shooting and specifications for water were regulated by two irrigation systems based on a mean-± standard error at P = 0.05 and graphically based on excel 2010.
RESULTS AND DISCUSSION
WUE's interaction with root systems
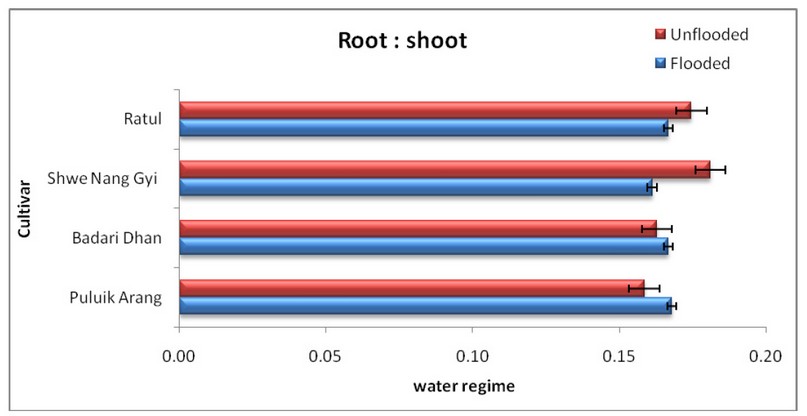
Figure.1 Mean± Standard Error of Root: Shoot ratio of four rice cultivars subjected to flooded and unflooded conditions. (Figure design depended (36) results at p < 0.05 according to Turkey’s test)
The high-water absorption potential in Figure.1 results in Puluik Arang are more excellent than Badari Dhan for the conservation of root: shoot ratio, while Badari Dhan was significant among the cultivars Ratul and Shwe Nang Gyi. In Puluik Arang, the results were outstanding. Puluik Arang showed no significant root variations under the unflooded condition: shooting between the cultivars (Badari Dhan, Shwe Nang Gyi and Ratul).
Such results confirm the physiologic and morphological characteristics of spark-and-rain cultivar Puluik Arang and Badari Dhan which are prone to unsafe conditions and are thus associated with water absorption and which therefore adds significant bio-mass renewability under a flooded environment. Research showed that genetic variation in root function was essential to better water intake under stressful conditions 50.
At root: the root ratio was reduced under the flood conditions compared with flooded areas, irrespective of the cultivar. Similar results have been documented in other experiments under soil moisture conditions and were generally limited even when water is mild or the soil is saturated 37; 38.
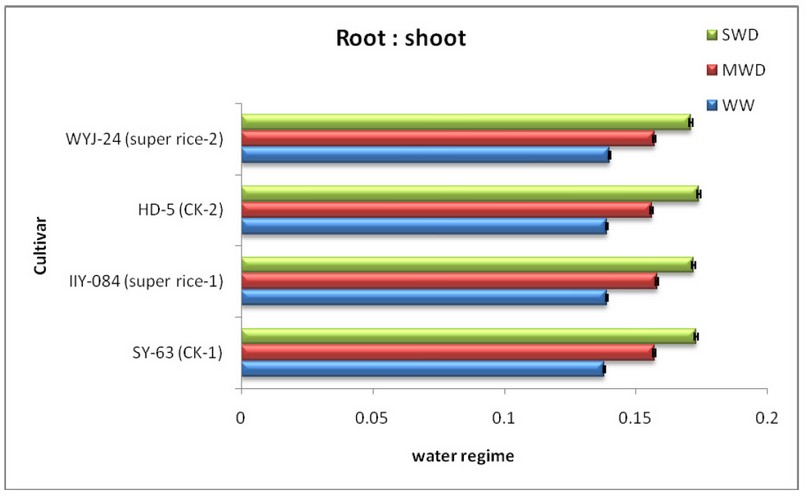
Figure.2 Mean± Standard Error of Root: Shoot ratio of four rice cultivars subjected to well-watered (WW), moderate water deficit (MWD), and severe water deficit (SWD) conditions. (Figure design depended on 41 results at p < 0.05 according to Turkey’s test)
The findings in figure 2 indicate the same actions as root: corresponding shooting ratio for both crops. When water surpluses rose, significantly below the SWD, moderately below MWD and lowest below the WW, the cultivars decreased considerably and no noticeable difference between the four varieties under WW.
The findings indicate either MWD or SWD: the root ratio decreased as water deficit increased to deal with soil water deficits and is better at growing rice cultivars. The root ratio decreased. No significant difference in moisture treatment between four cultivars: fire ratio showing the greater photosynthetic potential of the root and the snorkel, especially in soil deficits. The expectation is that small root biomass can sustain a strong biomass production over the field, while a deep root distribution will maximize land moisture capture and thus hold plant water in the high condition under dry conditions 39; 40.
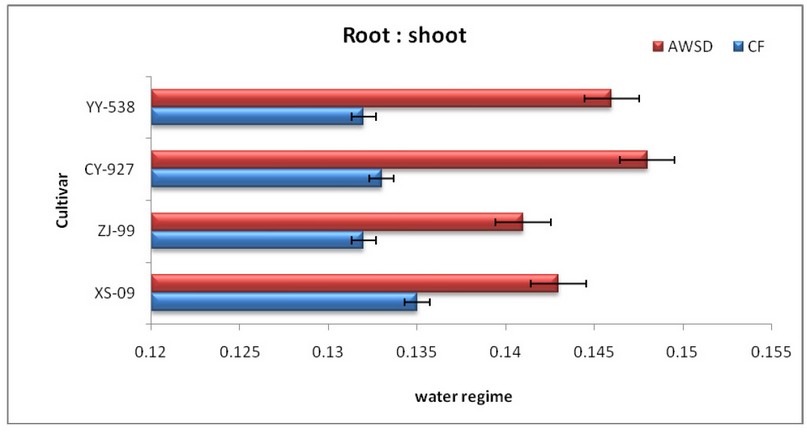
Figure. 3 Mean± Standard Error of Root: Shoot ratio of four rice cultivars subjected to alternate wetting and severe drying (AWSD) and constant flooding (CF) conditions.
(Figure design depended on 44 results at p < 0.05 according to Turkey’s test)
Results in figure 3 show that the AWSD irrigation system was highly significantly increased root: shoot ratio among cultivars relative to CF, with a more significant reduction in CY-927> YY-538> XS-09> ZI-99 varieties consecutively, but also better root: shoot ratio performance and highly under the CF irrigation regime compared with the XS-09 cultivars compared with other cultivars. Larger root: association of shoot with the deeper distribution of root and photosynthesis of plants with IJHR cultivar AWSD irrigation system. The experimental variations were due to changes in the hydrological component of the soil and the pacing of irrigated techniques. Better grains production and WUE for AWMD is mainly due to reduced vegetative residual growth and improved stability of canopy and increased root growth in hormone levels. Increases abscisic acid concentration and cytokinin levels usually during soil drying and decreased carbon transfer from tissue to vegetative grain during rewatering 37; 38.
NUE's interaction with root systems
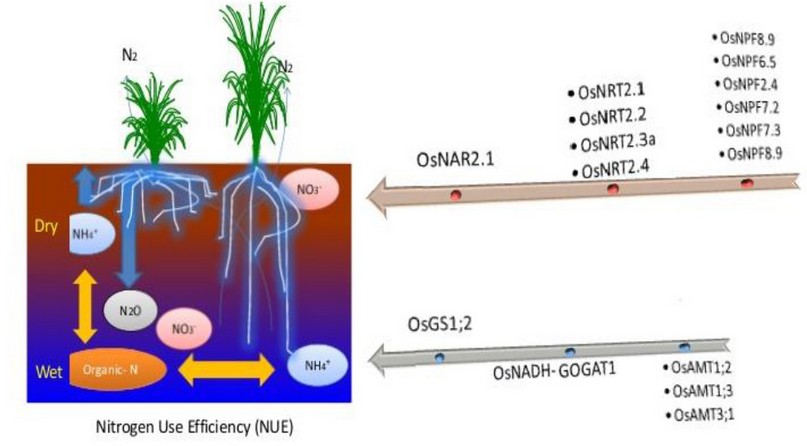
Figure 4. Schematic description of identified and predicted roles of NRT and NPF family rice nitrate carriers, AMT family ammonium carriers, and GOGAT nitrogen assimilation proteins 32; 46.
In figure 4 simulated, the analyze the root-shooting and root-soil relationships are the underlying role for higher seed production, the root-sourced hormonal roles in regulating crop development and growth, and the effect of soil moisture and nutrient distribution on root morphology and metabolism. The overall root productivity was significantly lower than in flooded plants due to the reduction in root abundance in the soil. Due to the significant reduction in the number of preventive plants, the role of fast root growth in soil water absorption and, hence, air perspiration survival, the weak ratio of root to shoot and disadvantaged root production in the surface layer of aerobic culture. The root morphology under upland rice included some portion of an ideotype to improve N capture for potential NO3- passage in the shaped profile at lower depths, but root appropriation varies unambiguously with soil conditions, supplement availability, and mechanical impedance. Upland rice's root morphology is an ideotype for optimizing N collection, extracting nitrate from lower root profiles, but its root distribution varies widely with the soil characteristics, fertilizer efficiency and mechanical impedance. NRT, NAR (OsNAR 2.1, 2.2), OsNRT, has functions for upregulating the transcription of N-use-related genes for both NO3 and NH4 +, and also for floral genes, for the most part of the NO3-transport families (OsNRT 2.1; 2.2, 2.3 and 2.4). NPF family members described to date were low-affinity NH4 transporters, with the especially that OsNPF family (8.9; 6.5; 2.4; 7.2 and 7.3) demonstrated dual-affinity nitrate transportation behavior associated with increased NO3- absorption and root-to-shoot transportation. OsNPF6.5, called an OsNPF8.9 putative mRNA splicing drug, has a major impact on both NUE and yield OsNPF8.9, expressed predominantly in root Skin and epidermis were cloned to cause N absorbance. Location of OsNPF4.1 in rice panicle elongation and OsNPF8.20 (OsPTR9) role in the absorption of NH4 +, lateral root development and decreased kernel yield were shown to work. The NUE-related QTLs and glutamine synthetase 1 (OsGS1;2), OsAMT(1;2, 1;3, and 3;1) and Glutamine oxoglutarate aminotransferase1 (OsNADH-GOGAT1) are affinity with NH4 transporters (32;43; 44;45). Here we noted that OsNRT (OsNPFs), a member of the rice (Oryza sativa) nitrate transporter family 1/peptide transporter, is involved in regulating N intake and yield, Gene over-expression in different cultivars appeared not to affect root size or WUE, being a good candidate gene for optimizing NUE and yield has been significantly improved, these conclusions mean that under stress conditions root uptake increases area uptake 48;49, root: shoot ratio has finally been correlated with grain uptake N.
CONCLUSION AND RECOMMENDED
NH4+ and NO3- is a major driving factor for growing crop yields, but with considerable flowering delays, prolonging the maturation and thus increasing the risk of yield losses. Therefore, characteristics that use high N levels without slowing ripening are highly desirable for plant breeding. The root morphology for upland and flood rice is part of an ideotype to optimize N aggregation, absorb nitrate at lower root profile depths, but the root distribution varies widely with soil capital, fertilizer quality, and irrigation system. Gene over-expression in different cultivars appeared not to affect yield or NUE or WUE, being a functional candidate gene for optimizing NUE and return has been significantly improved. In rice, these conclusions mean that under stress conditions root uptake increases area uptake, root: shoot ratio has finally been correlated with grain uptake N.
REFERENCES
1. Comas, L. H., Becker, S. R., Cruz, V. M. V., Byrne, P. F., Dierig, D. A. (2013) Root traits contributing to plant productivity under drought, Front. Plant Sci. 4:442.
2. Chen, L., Wang, G., Chen, P., Zhu, H., Wang, S., Ding, Y. (2018) Shoot-Root Communication Plays a Key Role in Physiological Alterations of Rice (Oryza sativa) Under Iron Deficiency, Front Plant Sci. 9:757.
3. Fageria, N. K, Moreira, A. (2011) The Role of Mineral Nutrition on Root Growth of Crop Plants. Advances in agronomy, 110(1):251-331.
4. Yamauchi, T., Colmer, T. D., Pedersen, O., Nakazono, M. (2018) Regulation of Root Traits for Internal Aeration and Tolerance to Soil Waterlogging-Flooding Stress, Plant Physiol., 176, 1118–1130.
5. Mishra, A. (2019) Morphological and Physiological Root Plasticity and Its Relationships with Shoot Growth of Rice with Water Regimes and Microbial Densities, IntechOpen.
6. Gu, D., Zhen, F., Hannaway, D. B., Zhu, Y., Liu, L., Cao, W. (2017) Quantitative classification of rice (Oryza sativa L.) root length and diameter using image analysis, PloS ONE 12: e0169968.
7. Menguer, P. K., Sperotto, R. A., Ricachenevsky, F. K. (2017) A walk on the wild side: oryza species as source for rice abiotic stress tolerance, Genet. Mol. Biol. 40(Suppl. 1), 238–252.
8. Jing, L., Rui, X., Chunchao, W., Lan, Q., Xiaoming, Z., Wensheng, W., Yingbin, D., Lizhen, Z., Yanyan, W., Yunlian, C., et al. (2018) A heading date QTL, qHD7.2, from wild rice (Oryza rufipogon) delays flowering and shortens panicle length under long-day conditions, Sci. Rep, 8: 2928.
9. Sandhu, N., Subedi, S. R., Singh, V. K., et al. (2019) Deciphering the genetic basis of root morphology, nutrient uptake, yield, and yield-related traits in rice under dry direct-seeded cultivation systems, Sci Rep, 9(1):9334.
10. Paez-Garcia, A., Motes, C. M., Scheible, W-R., Chen, R., Blancaflor, E. B., Monteros, M. J. (2015) Root traits and phenotyping strategies for plant improvement, Plants 4: 334–355.
11. Huang, S., Zhao, C., Zhang, Y., Wang, C. (2017) Nitrogen use efficiency in rice. In: Nitrogen in agriculture-updates,Amanullah and Shah Fahad. IntechOpen, London.
12. Wang, M.Y., Siddiqi, M.Y., Ruth, T.J., Glass, A. (1993) Ammonium uptake by rice roots (II. Kinetics of 13NH 4+ influx across the plasmalemma), Plant Physiol, 103, 1249–1258.
13. Yi, J., Gao, J., Zhang, W., Zhao, C., Wang, Y., Zhen, X. (2019) Differential Uptake and Utilization of Two Forms of Nitrogen in Japonica Rice Cultivars From North-Eastern China. Front Plant Sci, 10:1061.
14. Chen, G., Guo, S., Kronzucker, H.J., Shi, W. (2013) Nitrogen use efficiency (NUE) in rice links to NH4+toxicity and futile NH4+cycling in roots, Plant and Soil, 369:351–363.
15. Zhu, Y., Di, T., Xu, G., Chen, X., Zeng, H., Yan, F., Shen, Q. (2009) Adaptation of plasma membrane H+-ATPase of rice roots to low pH as related to ammonium nutrition, Plant, Cell and Environment, 32, 1428–1440.
16. Flam-Shepherd, R., Huynh, W.Q., Coskun, D., Hamam, A.M., Britto, D.T., Kronzucker, H.J. (2018) Membrane fluxes, bypass flows, and sodium stress in rice: The influence of silicon, J. Exp. Bot, 69, 1679–1692.
17. Britto, D.T., Siddiqi, M.Y., Glass, A.D.M., Kronzucker, H.J. (2001) Futile transmembrane NH4 + cycling: a cellular hypothesis to explain ammonium toxicity in plants, Proceedings of the National Academy of Sciences, USA, 98, 4255–4258.
18. Goyal, S. S., Tischner, R., Basra, A.S. (2005) Enhancing the efficiency of nitrogen utilization in plants, New York: Food Products Press.
19. Bloom, A.J., Sukrapanna, S.S., Warner, R.L., (1992) Root respiration associated with ammonium and nitrate absorption and assimilation by barley, Plant Physiology, 99, 1294–1301.
20. Ben-Noah, I., Friedman, S.P. (2018) Review and evaluation of root respiration and of natural and agricultural processes of soil aeration, Vadose Zone J, 17:170119.
21. Chauhan, B.S., Jabran, K., Mahajan, G., (Eds.), (2017) Rice Production Worldwide, Springer International Publishing, Cham.
22. Serraj, R., McNally, K.L., Slamet–Loedin, I., Kohli, A., Haefele, S.M., Atlin, G., Kumar, A., (2011) Drought resistance improvement in rice: an integrated genetic and resource management strategy, Plant Prod. Sci. 14, 1–14.
23. Rebolledo, M., Dingkuhn, M., Péré, P., Mc Nally, K., Luquet, D. (2012) Developmental dynamics and early growth vigour in rice. I Relationship between development rate and growth, J Agron Crop Sci (11 p.).
24. Smith, P., Martino, D., Cai, Z., Gwary, D., Janzen, H., Kumar, P., et al. (2008) Greenhouse gas mitigation in agriculture, Philos. Transac, 363, 789–813.
25. Liu, T., Q., et al (2015) Deep placement of nitrogen fertilizers reduces ammonia volatilization and increases nitrogen utilization efficiency in no-tillage paddy fields in central China, Field Crops Res, 184, 80–90.
26. Ali, J., Jewel, Z.A., Mahender, A., Anandan, A., Hernandez, J., Li, Z. (2018) Molecular genetics and breeding for nutrient use efficiency in rice, Int J Mol Sci. 19:1762.
27. Hirel, B.T., Tétu, P.J., Lea, F. (2011) DuboisImproving nitrogen use efficiency in crops for sustainable agriculture, Sustainability, 3, 1452-1485.
28. Yu, J., Zhen, X., Li, X., Li, N., Xu, F. (2019) Increased autophagy of rice can increase yield and nitrogen use efficiency (NUE), Front. Plant Sci, 10, 584.
29. Anis, G.B., Zhang, Y., Islam, A., et al. (2019) RDWN6XB, a major quantitative trait locus positively enhances root system architecture under nitrogen deficiency in rice, BMC Plant Biol. 19(1):12.
30. Zhang, Q.F. (2007) Strategies for developing green super rice, Proc Natl Acad Sci USA, 104:16402–9.
31. Yang, J.C., Zhang, H., Zhang, J.H. (2012) Root morphology and physiology in relation to the yield formation of rice, J. Integr. Agric, 11 (6), 920–926.
32. Obara, M., Kajiura, M., Fukuta, Y., Yano, M., Hayashi, M., Yamaya, T., Sato, T. (2001) Mapping of QTLs associated with cytosolic glutamine synthetase and NADH-glutamate in rice (Oryza sativa L.), Journal of Experimental Botany, 52: 1209-1217.
33. Kato, Y., Okami, M., Katsura, K. (2009) Yield potential and water use efficiency of aerobic rice (Oryza sativa L.) in Japan, Field Crops Res, 113:328–334.
34. Mcdonald, A. J., et al (2008) Stubborn facts: Still no evidence that the System of Rice Intensification out-yields best management practices (BMPs) beyond Madagascar
35. Peng, S., Tang, Q., Zou, Y. (2009) Current status and challenges of rice production in China, Plant Prod Sci, 12(1):3–8.
36. Matsunami, M., Matsunami, M., Kodama, I., Ogawa, A., Toyofuku, K., Ishikawa-Sakurai, J., Kokubun, M. (2016) Characterization of the morphological and physiological traits of rice cultivars with adaptation to unflooded condition during early vegetative growth, Plant Production Science, 19:1, 173-180.
37. Kano-Nakata, M., Inukai, Y., Wade, L.J., Siopongco, J.D. L.C, Yamauchi, A. (2011) Root development, water uptake, and shoot dry matter production under water deficit conditions in two CSSLs of rice: Functional roles of root plasticity, Plant Production Science, 14, 307–317.
38. Kato, Y., Okami, M. (2010) Root growth dynamics and stomatal behavior of rice (Oryza sativa L.) grown under aerobic and flooded conditions, Field Crops Research, 117, 9–17.
39. Ju, C.X., Buresh, R.J., Wang, Z.Q., Zhang, H., Liu, L.J., Yang, J.C., Zhang, J.H. (2015) Root and shoot traits for rice varieties with higher grain yield and higher nitrogen use efficiency at lower nitrogen rates application, Field Crops Research,175, 47–59.
40. Chu, G., Wang, Z,Q., Zhang, H., Yang, J.C., Zhang, J.H. (2016) Agronomic and physiological performance of rice under integrative crop management, Agronomy Journal, 108,117–128.
41. Zhou, Q., Ju, C.X., Wang, Z.Q., Zhang, H., Liu, L.J., Yang, J.C., Zhang, J.H. (2017) rain yield and water use efficiency of super rice under soil water deficit and alternate wetting and drying irrigation J. Integr. Agric., 16,1028-1043.
42. Zhang, H., Chen, T.T., Wang, Z.Q., Yang, J.C., Zhang, J.H. (2010) Involvement of cytokinins in the grain filling of rice under alternate wetting and drying irrigation. Journal of Experimental Botany, 61, 3719-3733.
43. Zhang, Y.N., Liu, M.J., Saiz, G., Dannenmann, M., Guo, L., Tao, Y.Y., Shi, J.C., Zuo, Q., Butterbach-Bahl, K., Li, G.Y., Lin, S. (2017) Enhancement of root systems improves productivity and sustainability in water saving ground cover rice production system, Field Crops Research, 213, 186-193.
44. Chu, G., Chen, T., Chen, S., Xu, C., Wang, D., Zhang, X. (2018) The effect of alternate wetting and severe drying irrigation on grain yield and water use efficiency ofIndica-japonica hybrid rice (Oryza sativa L.), Food and Energy Security, 7, Article e133.
45. Ogawa, S. (2016) Rice Root physiology work at CIAT: Identification of ideal root system to improve water and Nitrogen uptake under stress conditions. Presentation. International Center for Tropical Agriculture (CIAT).
46. Hamaoka, N.; Uchida, Y.; Tomita, M.; Kumagai, E.; Araki, T.; Ueno, O. Genetic variations in dry matter production, nitrogen uptake, and nitrogen use efficiency in the AA genome Oryza species grown under different nitrogen conditions. Plant Prod. Sci. 2013, 16, 107–116.
47. Atiyah.Ameerah.H. , El -Kaaby Ekhlas A.J. , Mouhamad, R. S., Raied. H. and AlAnny .Jenan A. (2017) In Vitro Influence of drought on some physiological parameters in callus induced from seeds of four Rice cultivars (Oryza sativa L.). Int. J. of Multidisciplinary and Current research, Vol.5:1000-1003.
48. Mouhamad, R. S., Mutlag LA, Al-Khateeb MT, Iqbal M, Nazir A, Ibrahim KM, Mussa RA, Jassam OH,(2017) Salinity tolerance at seedling stage for rice genotypes: In vitro analysis. Net J Agric Sci, 5: 114-120.
49. Mouhamad, R. S., Jaafar ZM, El–Kaaby EAJ, Iqbal M, Arif N (2018) Evaluation of Agronomic Traits and Inorganic Nutritional Composition of Rice Seed from IRSSTN Genotypes in Iraq. J Rice Res 6: 189.
50. Mouhamad, R. S., Ameerah H Atiyah, Naghum A Masamsh and Ibrahim B Razaq (2017) Organic composition of IRSSTN genotypes rice evaluated under Iraqi climate. JEZS 2017; 5(5): 1831-1837.
Received: 23 December 2019
Accepted: 20 January 2020
Raghad S. Mouhamad
1Department of Soil and Water Resources, Ministry Science and Technology, Baghdad, Iraq
Corresponding Author, e-mail: [email protected]