2022.07.04.54
Files > Volume 7 > Vol 7 No 4 2022
Previous / Index / Next
Current situation of snakebites envenomation in the Neotropics: Biotechnology, a versatile tool in the production of antivenoms.

Elizabeth Romo 1 , Marbel Torres 1,2,3
, Marbel Torres 1,2,3 , Sarah Martin-Solano 1,2,4,*
, Sarah Martin-Solano 1,2,4,* .
.
1 Carrera de Ingeniería en Biotecnología, Departamento de Ciencias de la Vida y la Agricultura, Universidad de las Fuerzas Armadas-ESPE, Sangolquí, Ecuador; [email protected];2 Grupo de Investigación en Sanidad Animal y Humana (GISAH), Carrera de Ingeniería en Biotecnología, Departamento de Ciencias de la Vida y la Agricultura, Universidad de las Fuerzas Armadas-ESPE, Sangolquí P.O. Box 171-5-231, Ecuador, [email protected]
3 Immunology and Virology Laboratory, Nanoscience and Nanotechnology Center, Universidad de las Fuerzas Armadas, ESPE, Sangolquí, Ecuador; [email protected];
4 Grupo de Investigación en Biodiversidad, Zoonosis y Salud Pública, Universidad Central del Ecuador.
* Correspondence: [email protected];
Available from: http://dx.doi.org/10.21931/RB/2022.07.04.54
ABSTRACT
Snakebite envenomation is a neglected tropical disease that affects millions of people around the world with a great impact on health and the economy. Unfortunately, public health programs do not include this kind of disease as a priority in their social programs. Cases of snakebite envenomations in the Neotropics are inaccurate due to inadequate disease management from medical records to the choice of treatments. Victims of snakebite envenomation are primarily found in impoverished agricultural areas where remote conditions limit the availability of antivenom. Antivenom serum is the only Food and Drug Administration-approved treatment used up to date. However, it has several disadvantages in terms of safety and effectiveness. This review provides a comprehensive insight dealing with the current epidemiological status of snakebites in the Neotropics and technologies employed in antivenom production. Also, modern biotechnological tools such as transcriptomic, proteomic, immunogenic, high-density peptide microarray and epitope mapping are highlighted for producing new-generation antivenom sera. These results allow us to propose strategic solutions in the Public Health Sector for managing this disease.
Keywords: antivenom, biotechnology, neglected tropical disease, omics, recombinant antibody.
INTRODUCTION
Neglected diseases occur in tropical and subtropical climates, specifically in rural areas where access to clean water, sanitary conditions, and medical care are limited1. They are caused by various pathogens such as viruses, fungi, bacteria, parasites and toxins, causing health, economic and social consequences. The term neglected is because these diseases are absent in most public health programs2 affecting many people generating disability3 and unemployment4. In addition, these diseases cause significant effects on the economy of developing countries due to the high cost of the treatments5,6.
The World Health Organization WHO, in its NTDs portfolio, included 17 snake-caused diseases. Snakebite diseases were previously not on the NTD list, but since 2017, public health strategies have been planned for prevention, control and treatment7. Snakebites envenomation is found in Latin America, Africa, Asia and Oceania, poor rural tropics populations8-11. These countries have an absence of public health policies 12 pointed to snakebites diseases, so; they have no access to health services1,13 and have a shortage of both medical supplies and trained human medical equipment.
More than 4 000 snake species worldwide, but only 250 are known of medical relevance2. Regions with the most incredible diversity of venomous snakes include Latin America and Asia12. Snakebite should be considered an NTD priority because it involves a wide snake diversity of species and, thus, a variety of toxins14. Snakes causing most ophidian accidents belong to the Viperidae and Elapidae families and the genera Bothrops and Micrurus5,15,16. Several risk factors, such as climate17 and ecology18, predispose to increased ophidian accidents19. Both rainy seasons and snake abundance5 cause a higher snakebites incidence5,20,21.
Generally, global data for snakebites are not accurate, showing variability mainly due to scarce and not representative epidemiological studies 5,19,21. Most hospital reports22 and surveys12 do not report important data such as incidence, mortality, and physical and psychological consequences suffered by patients23. According to Pach et al.21, five million snakebites are reported annually worldwide, two to three million results in poisonings and 80 000 to 130 000 people die from these diseases. In Latin America and the Caribbean, hospital reports indicate approximately 70 000 cases of snakebites per year, which may be underestimated24,25.
As a mega-diverse country, Ecuador has 40 poisonous snake species22, of which 17 are responsible for 99% of poisoning cases. The most significant number of patients are found in the Amazon region, followed by the coastal area and the Andean region26, which correlates with one of the studies of the geographic pattern of poisonous snakes22. Among the toxic snake families, the most representative is Viperidae and Elapidae 23,26.
Poisons are a set of proteins, peptides and enzymes that cause toxic effects20 in the pathology of snakebite envenomation. The toxic profiles of each venom vary according to the geographic location and snake taxonomy27,28, generating a wide range of local and systemic pathologies, including blindness29,30, necrosis31,32, paralysis33, respiratory34,35, renal36,37 and cardiac insufficiency38. The only specific treatment currently available is antivenom (Table 1) or antisera since its development in 189439,40. The traditional method for producing antivenoms is based on animal hyperimmunization with non-lethal venom doses with the following collection of large amounts of plasma11,41.
Almost all countries in the world have limited antivenom availability1,13,42,43 due to low interest in drug research and development, costs, safety, efficacy, and inefficient antivenom distribution5,13,21,23. However, several investigations are carried out with different types of antibodies44,45, using pharmacological molecules46,47 and innovative DNA immunization strategies48,49 to inhibit or reduce toxic effects.
This review aims to update the real situation of snakebites in the Neotropics from the epidemiological point of view and also to expose current biotechnological tools that could be implemented in the future to solve the drawbacks in the production and availability of antivenoms.
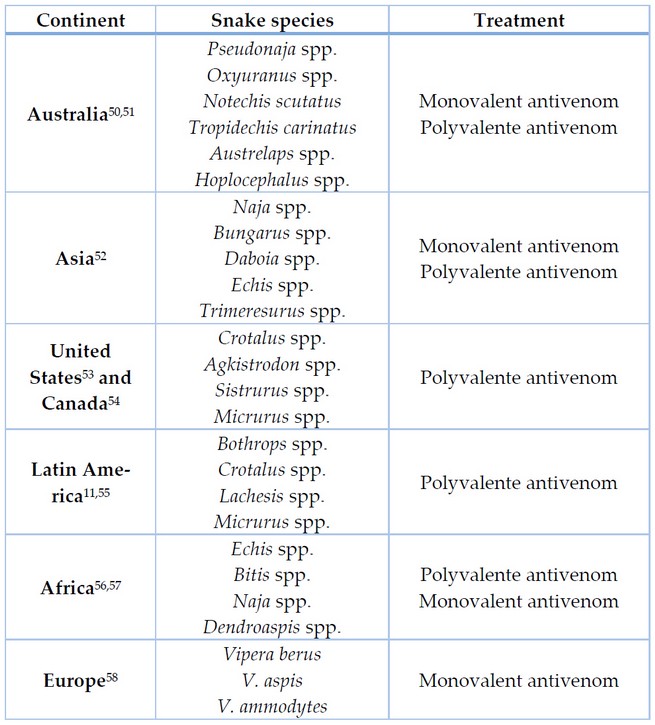
Table 1. Treatment for snakebite envenomation around the world.
Snakebite in the Neotropics: Ecological and Environmental Aspects
Ecosystem alteration, agriculture and environmental conditions modify the ecological patterns in the geographic distribution of the different snake species59. The highest incidence rate is concentrated in rural areas; urban spaces report snakebite cases60,61.
Venomous and non-venomous snakebites are the result of the interaction between humans and various snake species found in specific habitats such as jungles62, forests63,64, arid65,66 and urban areas67,68. Snakes, humans and the environment are the three components of this ecological interaction where one part can influence the other one while also affecting the third one. A circular dynamic characterizes this interaction, so it is necessary to know and relate these components. According to Guedes et al., 69 659 snake species were recorded in the Neotropics, where species richness and phylogenetic diversity are mainly concentrated in the Amazon region of Brazil, the Andean region of Colombia, Ecuador and Peru, and some Central America regions. On the other hand, few species are recorded in the Caribbean. In the Neotropics, the most abundant species are L. muta, Micrurus frontalis, B. jararaca, and Bothrops erythromelas69. These snakes are of medical and clinical importance in the Neotropics and belong to the Colubridae, Elapidae and Viperidae families70.
Agriculture causes geographic and ecological change in ecosystems, as forests are replaced by cultivated croplands71. Humans have been engaged in agricultural activities such as planting crops and raising animals since the Paleolithic era (12 000 to 5 000 years BP), which continued over the millennia to the present day72-74. Many people do this job and find themselves vulnerable to venomous snakebites75-77. According to Suazo-Ortuño et al.78 research on agricultural conversion, snakes are not susceptible to changes in their habitat; the species diversity in agricultural areas is not diminished, and snakebites risk is maintained. Several epidemiological studies determined a higher incidence in tropical rural areas where farming and grazing activities are carried out under unfavorable conditions8,14,19.
The most significant number of cases are recorded in rainy seasons associated with natural phenomena such as floods, hurricanes and cyclones79. In the Neotropics, the El Niño phenomenon causes heavy rains that increase the incidence of snakebites80. Snakes are ectoderm animals; temperature increases cause snakes to migrate to more temperate zones that are inhabited by humans81. Climate change then causes a geographic redistribution of snake species due to alterations in environmental temperature82-84.
Clinical findings, Social, and Economic Impact of Snakebite Envenomation
Venomous snakes can cause several local and systemic pathologies such as hemorrhage 85,86, necrosis 87,88 and renal failure89,90. Complications result in physical and psychological injuries91,92 in the short and long term, which can have an economic impact, including death.
Short-term sequelae occur immediately after the snakebite but can be controlled in a reasonable time. Among them, local hemorrhage93, anemia94, edema95, abscesses88 and bacterial infections96,97 are found. If these complications are not treated promptly and adequately, they can lead to systemic complications that can result in disability98.
Long-term sequelae appear approximately six months after snakebite envenomation and may persist for months or even years92,99. These sequelae are of the physical and psychological type, although the psychological effects have a late onset. For both, there is no follow-up for adequate treatment100. Among the physical sequelae, the most common are tissue injuries. Tissue necrotization can trigger compartment syndrome101, which must be addressed by surgical treatment. This procedure causes loss of tissue and skeletal muscle function3. In some cases, amputations must be performed that generate permanent disabilities102. The renal dysfunction developed in patients affected by ophidian accidents can be persistent and progress to acute and chronic renal dysfunction that strictly requires dialysis treatment103-105.
Psychological sequelae do not derive from the toxic effects of the envenomation. Still, they are the result of the traumatic process of the snakebite in which the patient suffers from the physical and economic consequences of the ophidian accident. Depression and post-traumatic disorder are the most reported effects affecting 25-45% and 43% of the patients evaluated, respectively, being the leading causes of morbidity106,107. Less common psychological effects are headaches, vertigo, hysteria, and delirium, but cognitive functions are not yet determined to be affected91,99. Long-term psychological sequelae also cause deterioration in the family and educational context. Patients present negative attitudes that prevent them from continuing their work, generating an economic and social impact107-109. Timely treatment would improve the life quality of snakebite victims. A first aid intervention, cognitive and behavioral psychotherapy allows for to reduction of psychological and psychiatric symptoms92,110.
In addition to the physical and psychological sequelae, the economic consequences are aggravated by the absence of rehabilitation after the ophitic accident. This fact hinders labor insertion. Most of the time, the rehabilitation cost must be assumed by the patient itself, who usually does not have the resources to do so. The family economy in areas with a high rate of snakebites is categorized as impoverished rural areas12,111.
Agricultural activity is the livelihood of these communities, being limited by the economic expense of the sequelae they suffer. According to these situations, conditions for a worthy life are reduced112-114. Finally, the economic impact affects the nuclear family and negatively affects local and national productivity115.
Unfortunately, some case studies do not detail the patient's conditions under which they die due to snakebites116. This fact makes it difficult to show how high a priority this disease is. Lizarazo et al.117 reported a case of a farmer who suffered a snakebite caused by the B. asper snake that produced a cerebral hemorrhage. Unfortunately, the antivenom administration was late; he presented a multiorgan failure and died. Hospital reports are vital in managing this neglected disease, so standardized processes should be implemented in public health centers.
Global and Regional Snakebites Burden
Global and regional snakebite envenomation burden has not been accurately determined because of scarce information and studies estimating the incidence and mortality of SBE 118. Existing data are based on hospitals' epidemiological reports that do not provide evidence of the true SBE burden24. Collecting information on snakebites and envenomations is difficult because most victims live in rural and remote areas with limited access to health services55. In addition, people in rural communities prefer to treat themselves with traditional methods and do not go to hospitals1.
In 1954 an estimated 500 000 poisonings were estimated119, and in 1998 the estimate increased to 5 million snakebites per year1,120. According to Gutiérrez et al.121, the snakebite burden has an estimated 2.5 million bites per year, concentrated in South Asia, sub-Saharan Africa and Latin America. Through a regional comparison, it has been possible to determine the global level and the regions most affected by snakebite envenomation. The annual envenomation cases vary from region to region: Europe, where non-venomous snakes mainly cause snakebites, reported 8000 cases; North America 5 000 to 10 000 patients; the Middle East 15 000 to 40 000, Africa 43 000 to 1 000 000, Asia 121 000 to 2 000 000, Australia 10 000, Oceania 10 000 to 500 000 and Latin America 60 000 to 300 000 1,120-122 respectively.
Snakebite occurs in different geographical environments, whose social, economic and ecological factors may be similar, allowing the development of social and technological strategies to cope with this disease in the context of public health. Gutiérrez et al.123 evaluated the snakebite envenomation situation in Costa Rica, Nigeria and Sri Lanka, the most affected regions worldwide. The annual SBE burden is similar in Nigeria, with 43 000 reported cases, and in Sri Lanka, 40 000 reported cases, while in Costa Rica, case reports were much lower, reporting just only 500 cases.
The scientific community in Neotropical countries has conducted several epidemiological studies. The results may overlap with other existing ones, and some countries do not publish any information24. In the Neotropics, the countries with the most scientific publications on snakebite envenomation are Costa Rica, Colombia, Ecuador, Argentina and French Guiana1.
The lack of data reliability and accuracy is due to deficient information systems in the different countries and because victims from rural areas prefer to use traditional treatment methods based on medicinal plants124,125. In Latin America, Chippaux5 reported around 60 000 cases between 2014 and 2016 of snakebites per year and just 370 deaths. In another study, Kasturiratne et al.1 made epidemiological estimates of SBE obtaining 115 000 cases of snakebites and 2 000 deaths from 1985 to 2007.
Variations in epidemiological indicators such as burden, incidence, prevalence and mortality are due to the influence of environmental and anthropogenic factors. Also, the period and geographical area of epidemiological evaluation and the El Niño current have different effects according to geographical location126. The countries with the highest incidence of around 100 000 cases of SBE per inhabitant in the Neotropics, according to the official health reports of each country, are Panama with 55.8, French Guiana with 21.1, Venezuela with 18.9, Costa Rica with 15.0 and Brazil with 13.45. Values of SBE burden vary among scientific publications and may be over or underestimated. The annual incidence of snakebites worldwide is about 6.2 per 100,000 inhabitants, while mortality is 0.04 per 100,000 inhabitants5.
Antivenom Production: Current Status in the Neotropics
Antivenom production in the Neotropics dates back to the beginning of the 20th century at the Butantan Institute in Brazil in 1901, considered one of the pioneering laboratories in the region118,127 together with the Clodomiro Picado Institute in Costa Rica, founded in 1970128. In 2014, countries such as Mexico, Costa Rica, Venezuela, Colombia, Ecuador, Peru, Bolivia, Brazil and Argentina were antivenom producers with laboratories in Public Institutions129,130. Brazil, Mexico and Costa Rica were able to satisfy the antivenom demand at the national level and even cover the regional and global market118,131-134.
In other cases, where antivenom needs are not supplied, it must be imported from other countries in the region and even from other continents, as in the case of Martinique and Saint Lucia, Caribbean islands, which import antivenom from France and the United States, respectively118,135. In the case of Ecuador, antivenom production was local at the former antivenom producer "Instituto de Higiene y Medicina Tropical, Leopoldo Izquieta Perez", which operated up to 2012 and was closed due to deficiencies in the production processes13. At the beginning of 2022, the National Institute of Public Health Research INSPI signed a cooperative agreement with the Regional Amazonic University IKIAM to implement a research project to optimize the experimental production of effective antiophidic sera in Ecuador. This project promotes a change in public health perception and in the snakebite envenomation victims' lives.
In addition to the importance of producing antivenoms to reduce the SBE burden, they should be validated through pre-clinical and clinical trials to determine their effectiveness against various venoms. This becomes in an essential task due to the diversity of snake species in the Neotropics. Several clinical studies have been conducted with antivenoms from the region to treat endemic snake envenomation136 and have successfully reduced the envenomation signs developed in the victims137-139. The results of these assays indicate the existence of cross-reactivity106,130,133,140,141among antivenoms and toxins affecting patients, while others cannot neutralize heterologous venoms142. Antivenoms produced commercially in the Neotropics at a laboratory scale are mostly derived from equine serum and neutralize venoms of the genus: Bothrops143-145, Crotalus146-148, Micrurus149-151, Lachesis140,152.
Currently, there are only three antivenoms approved by the Food and Drug Administration FDA for exclusive use in the United States: Antivenin® Wyeth153, CroFab®154 and Anavip®155. In the Neotropics, only the antivenom Antivipmyn®, manufactured by Bioclon Laboratories of Mexico, was recognized by the FDA as an orphan drug156. These orphan drugs are intended to treat diseases affecting a small number of people, less than 200,000157. However, the FDA-approved drugs derived from snake venoms such as Captopril and Batroxobin158, obtained from the venom of B. jararaca, and the latter from B. moojeni and B. atrox32,159. These drugs have proved biomedical applications.
The efforts of antivenom research and production are evident in the published scientific literature; however, there are drawbacks related to the heterogeneity in the used production technologies, the quality and innovation of the pharmaceutical products obtained and the production volumes129,160,161. These variables cannot be analyzed due to the absence of updated and reliable epidemiological information that would allow having a base-line and thus supply the antivenom requirements. In addition, antivenom production in Latin America during 2020 was reduced as a consequence of the COVID-19 pandemic caused by SARS-CoV-2. Medical supplies and research were mainly focused on developing therapies and diagnostic kits to cope with COVID-19 health emergencies162.
Snakebite in Ecuador
Epidemiological studies in Ecuador are limited62,163. No data has existed in the country's health system records in the last three years. From 2014 to 2018, the Public Health Ministry of Ecuador, through the Epidemiological Gazette, recorded 7 714 cases of snakebites in the country, with an average incidence of 9.37 / 100 000 inhabitants and an annual incidence of 9.0, 11.3, 10.4, 8.6 and 7.6 respectively164. In the Chippaux5 study, incidence and mortality rates of 9.5 and 0.057 cases per 100,000 inhabitants were estimated in 2014-2015. In another study, Ecuador reported 9.8 cases per 100,000 inhabitants that resulted in 0.06 deaths per 100,000 inhabitants each year in 1998-200780. The annual incidence of snakebites in Ecuador does not have significant variations even though the Spatiotemporal analysis differs, 11.5 /100 000 inhabitants80,7.7-11/100 000 inhabitants165 and 9.5 / 100 000 inhabitants5.
Morona Santiago and Manabí are the provinces that register the highest number of snakebite cases. However, it is observed that in both the coastal and Amazonic regions were found a more significant number of cases164. Snakebites are distributed geographically in the Coastal region (56-58%), Andean region (5-33%) and Amazonic region (11-37%)80. Studies conducted in indigenous communities indicate that the highest incidence of snakebites occurs in the Amazon region142,166-168, associated with the distribution of snake species in the country22,26. Among the species causing snakebites in the country is the genus: Bothrops, Bothriopsis, Lachesis y Micrurus79 and the species: B. atrox, B. asper, L. muta, B. bilineata y Bothrops lojanus26,168. Information obtained from available epidemiological studies indicates that those mainly affected are agricultural workers in rural areas80. Heavy rains such as the El Niño Phenomenon in January to June increase the incidence of snakebite cases throughout the country80.
Biotechnological Approach to Snakebite Therapy
Drawbacks of available Antivenoms
Antivenom therapy is the most widely used therapy to treat several pathologies, including snakebite envenomation, by neutralizing the venom proteins169. Traditional antivenom production has several drawbacks affecting their safety and efficacy, such as the venom complexity, adverse reactions and production costs. Venom composition varies by snake diversity69, geographic distribution170,171, ontogenetic172 variations and snake growth stage173. Toxins that make up snake venom have a great molecular and biological diversity, including protein and peptide content; non-protein components such as carbohydrates, lipids, amino acids, nucleosides, amines and metal ions174. The main molecules of medical importance are grouped into dominant and secondary proteins: phospholipases, metalloproteinases, serine proteinases, three-finger toxins, hyaluronidases, and myotoxins, among others (Table 2)175. These factors limit the antivenom's neutralization spectrum, although several studies in the Neotropics indicate a good neutralization capacity of heterologous venoms, as mentioned in the previous sections.

Table 2. Main snake venom toxins, physiological effects and its 3D-structure
Antivenoms are obtained from animal immunization, such as horses184,185, donkeys 186, and sheep187,188, hence the name heterologous antivenoms189,190. Antivenoms currently present several drawbacks, as 59% of patients treated with this therapy develop adverse events and side effects191. Early anaphylactic reactions cause headaches, vomiting, fever, and urticaria192,193 and may or may not be an IgE-mediated response194. Antivenom can be composed of complete IgG, antibody fractions (Fab or (Fab')2) and other serum proteins of animal origin that can also cause adverse reactions. In addition, many antibodies do not neutralize the target antigen195. Different types of antibodies are involved in developing late anaphylactic adverse reactions, such as human anti-horse antibodies named heterophile. These antibodies form an immune complex deposited in the target tissues causing inflammation known as serum sickness196,197.
Purification and enzymatic digestion techniques are being employed to improve the quality and safety of the drug product while reducing the effects of adverse reactions198. Antivenom formulations with antibody fragments maintain neutralizing capacity and minimize adverse effects137,199. This antibody-based production technique currently employed has several disadvantages due to the costs of animal maintenance and antibody purification techniques. These facts limit the reproducibility of this technology14,189.
Based on this background, we expose the use of biotechnological tools with an innovative approach to improve the neutralization of venoms, increasing their efficacy and improving production yields. It is essential to consider biotechnology as a solution to the shortage of antivenoms in Latin America and the world since production costs can be reduced, and the pharmaceutical market in the production of antivenoms will be empowered.
Recombinant Antibody Technology
The first recombinant antibodies were developed by Georges Kohler and Cesar Milstein in 1975 using the hybridoma technology200. Recombinant antibody technology since then has become a potential therapy for snakebite envenomation. They are more effective because of their neutralizing capacity and reduced side effects such as anaphylactic reactions compared to animal serum-based antivenoms198. Using this technology, several antibody formats have been obtained in chimeric and humanized versions, whose therapeutic use is approved by the FDA in treating several diseases201,202.
The hybridoma technology, nevertheless, has several disadvantages. The main one is related to the development of human anti-mouse antibodies that cause allergic reactions and decrease the lifetime of the therapeutic antibody203; however, this is still the most widely used technique for antibody production. Production of antibodies involves an immunological process in which the antigen undergoes proteolytic degradation, so the derived antibodies will not distinguish the antigen in its initial form204. Therefore, researchers work to find other technologies that solve these drawbacks and do not activate the complement system.
Antibodies for therapeutic purposes like IgG have a longer life and are more permeable205. Recombinant antibodies have various formats in the structure; they are assembled according to different combinations of heavy and light chains206. The single-chain variable fragment (scFv) and fragment antigen-binding137 are the most commonly used formats because of their high affinity to the antigen, structural stability and shorter generation207.
Currently, several studies demonstrate the efficacy and therapeutic potential in neutralizing various snake venoms using recombinant antibodies of multiple formats, e.g., camelids nanobodies against the poison of B. atrox208 and C. durissus terrificus209; scFv against the venom of Bothrops pauloensis210 and L. muta 211. Recombinant antibodies, such as scFv and nanobodies, are FDA approved for treating the diseases, except for the treatment of snakebite envenomation212. In addition, the FDA and European Medicines Agency EMA has not approved the production of recombinant antibodies in E. coli189.
The isolation of therapeutically effective antibodies has presented low yields due to the high purification costs using traditional methods213-216. Smith217, in 1985 developed the phage display technique (PDT) that is independent of an immune system, making this technique the most selected since it does not generate immunogenicity in patients218. The PDT, as shown in Figure 1, consists of an in vitro phenotypic selection of antibodies expressed with the fusion proteins of M13 filamentous phages. At the same time, a genotypic selection is needed because the various genes encoding the antibodies of interest are found inside the phages219,220. The antibody selection and enrichment process are performed by affinity to the molecules of interest, in this case, the venom toxins218,221. This technology's advantages focus on controlling the process conditions such as antigen selection, immobilization, design of the antibody libraries, and binding and washing needs. In addition, it is a faster and low-cost technique compared to hybridomas production204. It is good to stand out that antibody production against snake venom toxins has only reached the laboratory scale; research is still ongoing for their optimization and further scaling up.
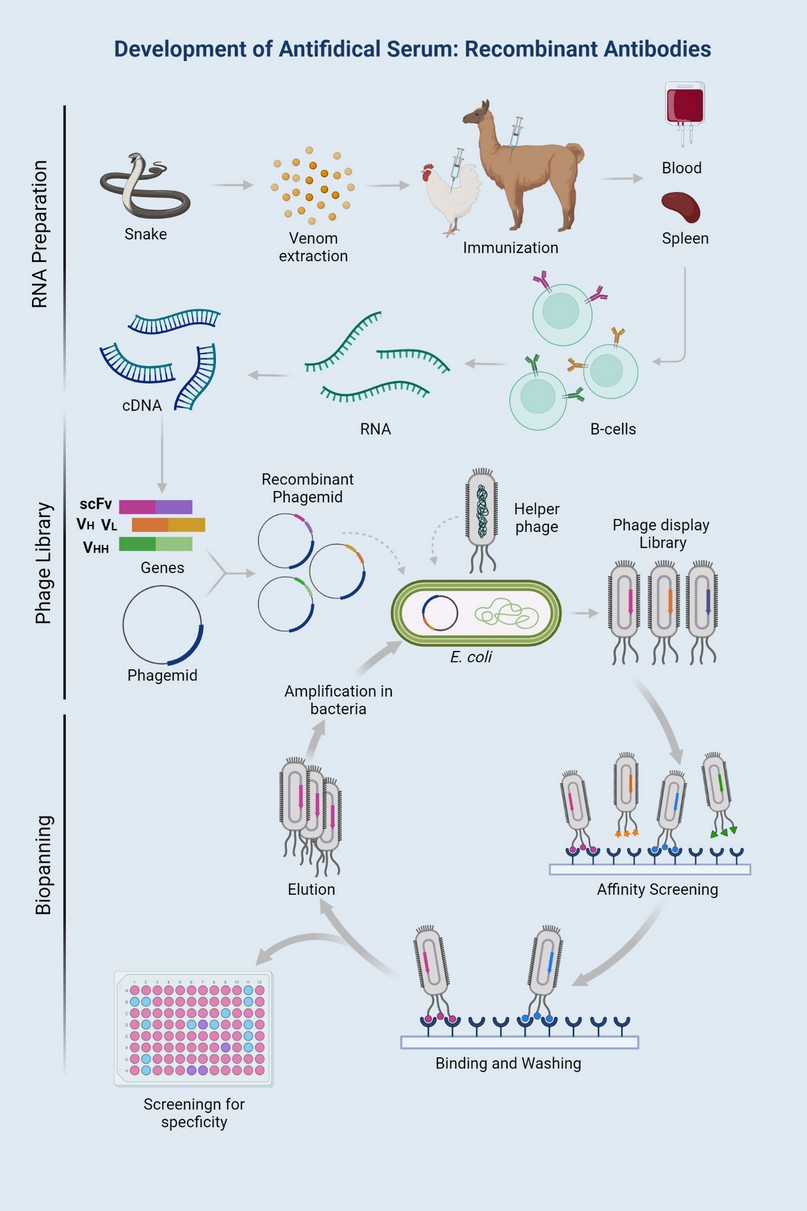
Figure 1. Phage display for selection of antibodies against snake antivenom. Image created using BioRender (https://biorender.com/).
The next stage of affinity antibody selection is heterologous expression. There are several expression systems ranging from bacteria, yeast, insects, and plants to mammals, each with advantages and disadvantages222. Bacteria such as Escherichia coli 223,224and Bacillus subtilis225,226 have been employed as factories to produce heterologous proteins for therapeutic purposes because their genome is characterized and genetic manipulation is simple; they have rapid growth and bioprocesses have enabled large-scale production at low cost167,227. Castro et al.228 produced a recombinant antibody, scFv, that neutralizes the BaP1 metalloprotein from Bothrops asper snake venom by expressing the antibody in a bacterial system using Escherichia coli as host (Figure 2). After protein extraction and purification, the yield was 280 ug of scFV per liter of bacterial culture. The drawbacks of these systems are due to the absence of post-translational modifications and a poor excretion system, as the stability of the proteins depends on the oxidative environment where it is secreted. In addition, inclusion bodies can be formed, hindering antibody purification229-231. Strategies to optimize antibody production in E. coli are listed in Table 3.
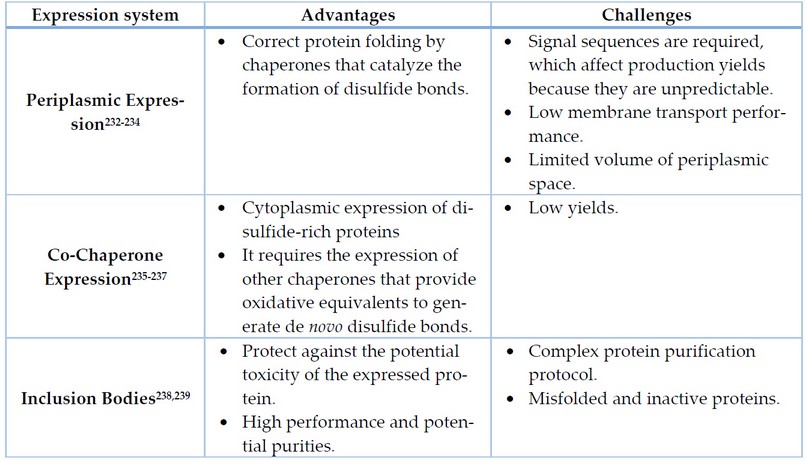
Table 3. Strategies for the expression of disulfide-rich proteins in E. coli.
The most commonly used yeasts for these purposes are Pichia pastoris240-242 and Saccharomyces cerevisiae243,244. Both are easy to grow, perform post-translational modifications such us disulfide-bonded and protein glycosylation, have a high growth rate and protein secretion levels are very high. Contrary to bacterial expression systems, protein secretion in yeasts constitutes a great productive advantage since the secreted proteins are harvested relatively quickly from the culture medium, so downstream processes are cheaper 242,245. Pichia pastoris is also the most widely used yeast strain due to its ease of industrial scaling. It reduces costs and minimizes equipment used for implementing pilot or industrial bioreactors246. Yeast expression systems are used to produce recombinant antibodies and proteins with inhibitory action against venom toxins (Figure 2). The antimyotoxic protein DM64, which acts against phospholipases A2 of Bothrops asper venom247, was successfully produced by a recombinant Pichia pastoris.
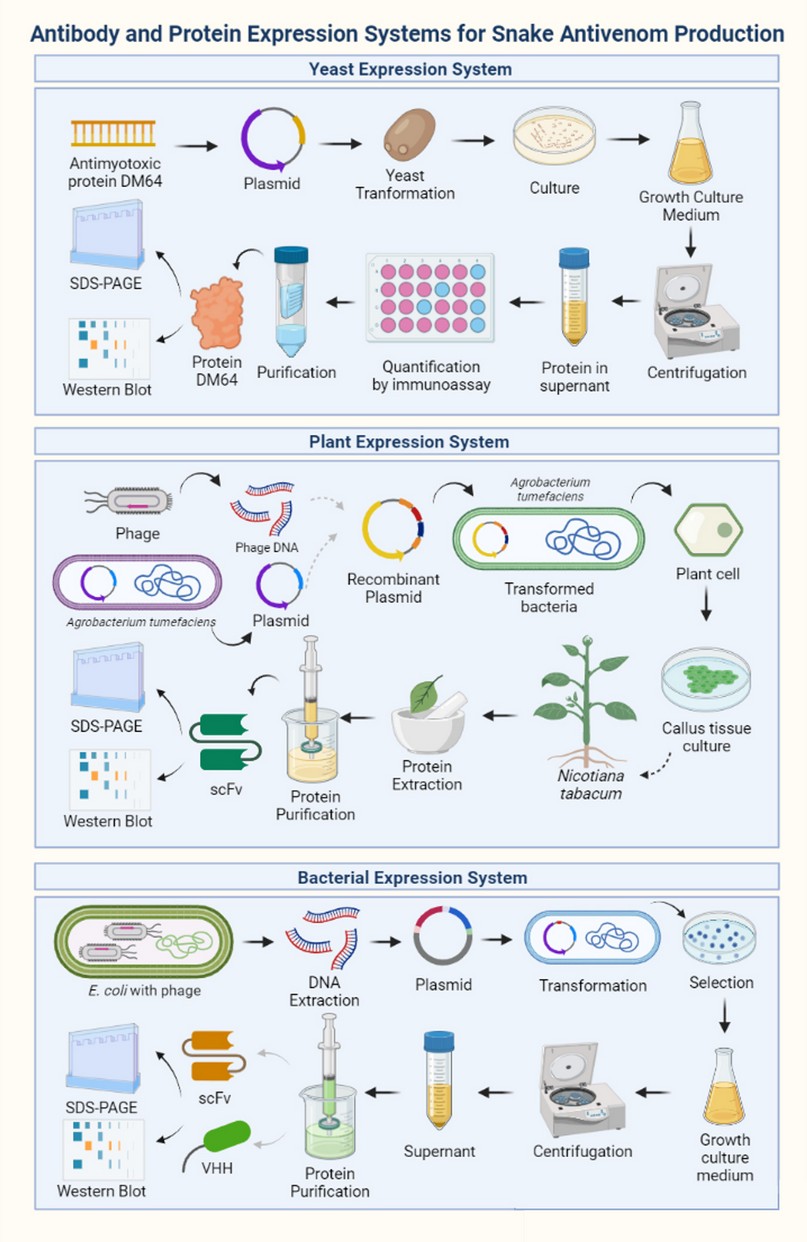
Figure 2. Protein and recombinant antibody expression systems for the production of snake antivenoms. Image created using BioRender (https://biorender.com/).
Mammalian cells are commonly used to produce biopharmaceuticals, antibodies and active protein248. Antibody production in mammalian cell systems is mainly selected by its ability to carry out post-translational modifications that maintain antibody stability so a correct protein function. However, expensive culture media due to nutrient requirements and high contamination rates limit this technology. In addition, yields are low and the slow production time increases costs236,249.
Laustsen et al.251 and Jenkins and Laustsen250 estimated the cost of large-scale production of antibodies in the Chinese Hamster Ovary (CHO) cell expression systems using a fed-batch fermentation strategy. The production cost ranged from 20 to 250 USD for these pharmaceutical products. On the other hand, plasma-derived antivenom production is around a thousand dollars. Currently, no studies are estimating the cost of new screening and expression technologies applied to large-scale antivenom production.
An alternative system to those described above includes insect cells advantageously as production hosts. This expression system can use chaperones for correct protein folding and own key metabolic pathways to carry out post-translational modifications, such as acetylation or glycosylation251,252. The system works with the baculovirus expression vector253. Insect cells are used as hosts to a greater extent for toxins production used as immunogens and for different in vitro toxicity assays. This technology is more complex but with high throughput and reproducibility at low costs254.
Finally, we have plant-based expression systems. To date, several types of toxin antibodies have been produced experimentally255-258. Plants are considered biofactories because of the amount of biomass they generate, allowing large-scale production. They are low-cost and not susceptible to contamination. Nevertheless, even though the initial steps of N-glycosylation and N-glycan processing are highly conserved between plants, mammals and yeast, N-glycosylation patterns differ between them259.
Antibody expression titers in plants are low, so approaches for expression improvement have pointed to expression cassette design, plant and tissue selection and plant material extraction techniques260. There are few studies of antibody expression in plants (Figure 2). One reported the extraction and purification from Nicotiana tabacum leaves of scFv against B. pauloensis venom210.
Although different techniques currently produce monoclonal antibodies, other biotechnological alternatives can be employed to enable regional and global scale of antivenoms production.
Omics for the production of Antivenoms
Omics enable innovation in the health sector to broaden the understanding of physiological processes of pathologies involving various molecules such as nucleic acids and proteins261 so, facilitating effective diagnosis and treatment262. Omics tools such as proteomics and transcriptomics are a fundamental axis in the design and production of antivenoms (Figure 3) as they are part of the preclinical evaluation and improvement of antivenom efficacy170,263.
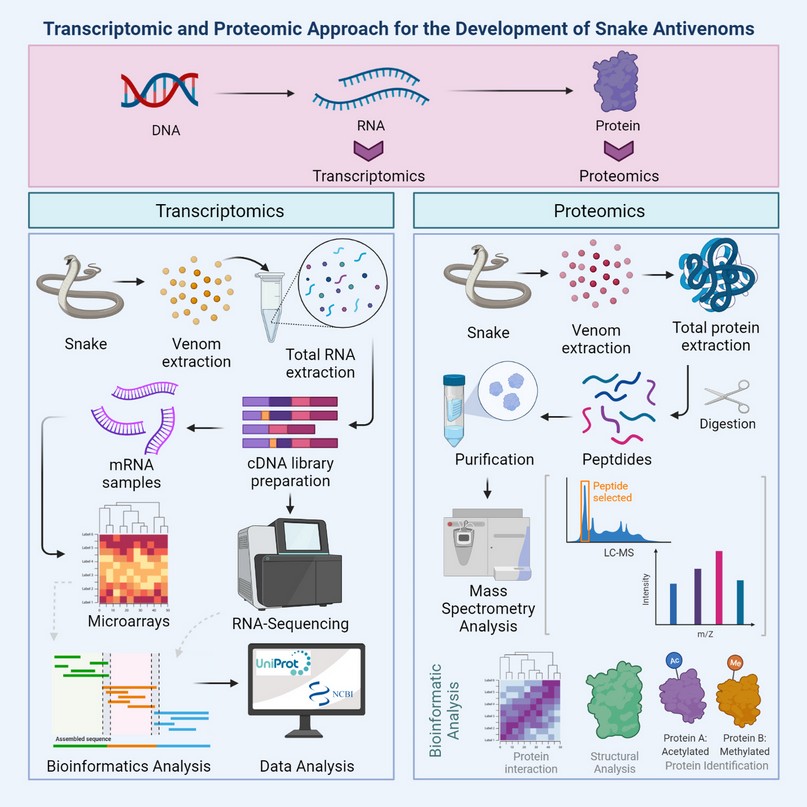
Figure 3. Transcriptomic and Proteomic Approach for the Development of Snake Antivenoms.
These technologies must comply with the good manufacturing practices detailed in the WHO Guidelines for the industrial production of antivenoms43,264 to ensure the quality and safety of the pharmaceutical product265. Proteomic and transcriptomic guide researchers to understand the biochemical and toxicological variations in venoms to antivenoms265 response. Omics tools should be included in antivenom production processes to validate the safety and quality of the bioproduct266.
Transcriptomics
Transcriptomics studies genome-encoded RNA transcripts such as mRNA, rRNA, tRNA, miRNA, and non-coding RNA267. The mRNA is required for protein synthesis, and its abundance indicates the presence of a target gene. The transcriptome is subject to change due to time, environmental and physiological conditions268. Transcriptomics gives information on RNA diversity, transcriptional units, splicing mechanisms, post-transcriptional modifications and information of gene expression, regulation and signaling267. The transcriptomics workflow is depicted in Figure 3.
Transcriptomic studies of venoms and venom glands of some snakes from the Neotropics were carried out by Rodrigues et al.269. He compared the transcriptomic profiles of the venom and venom gland of Bothrops pauloensis, finding qualitative variations and low concordance with the proteomic profiles. Ontogenetic changes affect venom composition; in young species of B. jararaca there is a greater diversity of toxin precursors and elevated amounts of metalloproteinases compared to adult species173. The analysis of the ontogenetic factor is fundamental in the production of antivenom since the efficacy in neutralizing envenomations caused by juvenile species may be limited. Freitas-de-Sousa et al.270 evaluated the environmental effect in captive and wild species of de B. atrox, the composition of the venoms does not present significant quantitative differences, thus, supporting the use of venoms from captive species for the production of the antivenom.
The use of transcriptomics as a tool for toxin discovery has displayed good results. In the venom gland of Bothrops moojeni, new toxins have been discovered, and amino acid sequences of unreported toxins have been obtained. These findings promise to know the function of new toxins and to design effective and neutralizing antivenoms271. Transcriptomics is used to know the complexity and composition of snake venoms and to evaluate toxins' immunogenicity at the molecular level, specificity and affinity for epitopes. The study of phospholipase A2 and three-finger toxin from Micurus nigrocinctus venom presented low immunogenicity272.
Proteomics
Proteins are expressed in cells and perform cellular processes related to biological functions. Proteomics studies the entire set of proteins in a cell or organism273. It is characterized by being dynamic and influenced by time, space, environment and cellular modifications such as post-translational modifications274,275.
Proteomics has several approaches to obtaining information about protein structure and functionality, including cellular expression, modifications, interactions and signaling275. The study of proteomics is essential if we consider proteins as gene products since proteins determine the phenotype. Genomics is static; the expression level of a gene will not always correlate with protein levels276 due to post-transcriptional and post-translational modifications. The study of snake venom proteins is also known as venomics. Both venomics and peptidomics allow understanding of the biological processes in envenomation, development of new therapeutics and potential pharmacological applications of snake venom toxins277. Proteomics follows two main experimental approaches for its study: gel-based and mass spectrometry. The mass spectrometry approach is divided into two modalities: Bottom-up or Shotgun proteomics, where proteins undergo enzymatic digestion, and top-down analysis employs intact protein278. The proteomics workflow is depicted in Figure 3.
Venom proteomics studies showed variations in the composition and functionality of toxins by several factors. A phylogeographic approach with proteomic tools has determined the venom phenotypes of snake species belonging to the Micrurus; the geographic distribution of venomous snakes and evolutionary mechanisms are very influential factors279. There are interspecific, intraspecific, qualitative and quantitative variabilities of snake venoms under different environmental conditions. Oliveira et al.165 evaluated the proteomic profiles of 22 individuals of the C. durissus terrificus, new venom components were found with various enzymatic activities that cause other immunological and biochemical effects affecting antivenom production. Snake species, such as B. atrox and B. jararaca, develop a response to adapting environments that can produce several venoms-protein isoforms at the molecular level with different biological activities, complexity and enzymatic activity, which limits antivenom efficacy280,281.
Immunogenomics
Immunogenomics is an essential tool for antivenoms development since epitopes mapping antibodies capable of recognizing these antigenic sites can be designed. This fact increases the neutralization capacity of antivenoms282. This tool, also known as antivenomics, allows identifying the recognition of certain immunogens by antibodies, a key factor in the clinical efficacy of antivenom in snakebite envenomation283.
The antivenoms produced at Instituto Butantan and Instituto Clodomiro Picado present efficient neutralization of the venom of B. atrox and Bothrops erythromelas species in the northern region of South America and Brazil, respectively170,284. The commercial antivenom Antivipmyn Tri produced in Mexico by Instituto Bioclón exhibits immunoreactivity of C. durissus cumanensis venom in Colombia285.
Toxins used to determine antibody responsiveness are also produced without the need for host cells. Protein synthesis is performed with the necessary components: dNTPs, amino acids, ATP, GTP, biological machinery that includes ribosomes, tRNA, RNA polymerases, initiation and elongation factors and a motor-like plasmids DNA carrying the correct information for transcription, translation and accurate folding in vitro. The main drawback of cell-free production is the low yield of proteins and their poor stability286,287.
Bioinformatics, the best omics ally
The results obtained from these omics tools are massive, so computational tools are needed to facilitate data analysis288,289. Bioinformatics, through the use of algorithms and computational strategies, develops methods to analyze biological data, which include: data organization and curation, processing, annotation, statistical analysis, prediction and simulation290. The most commonly employed bioinformatics analyses are listed in Table 4.
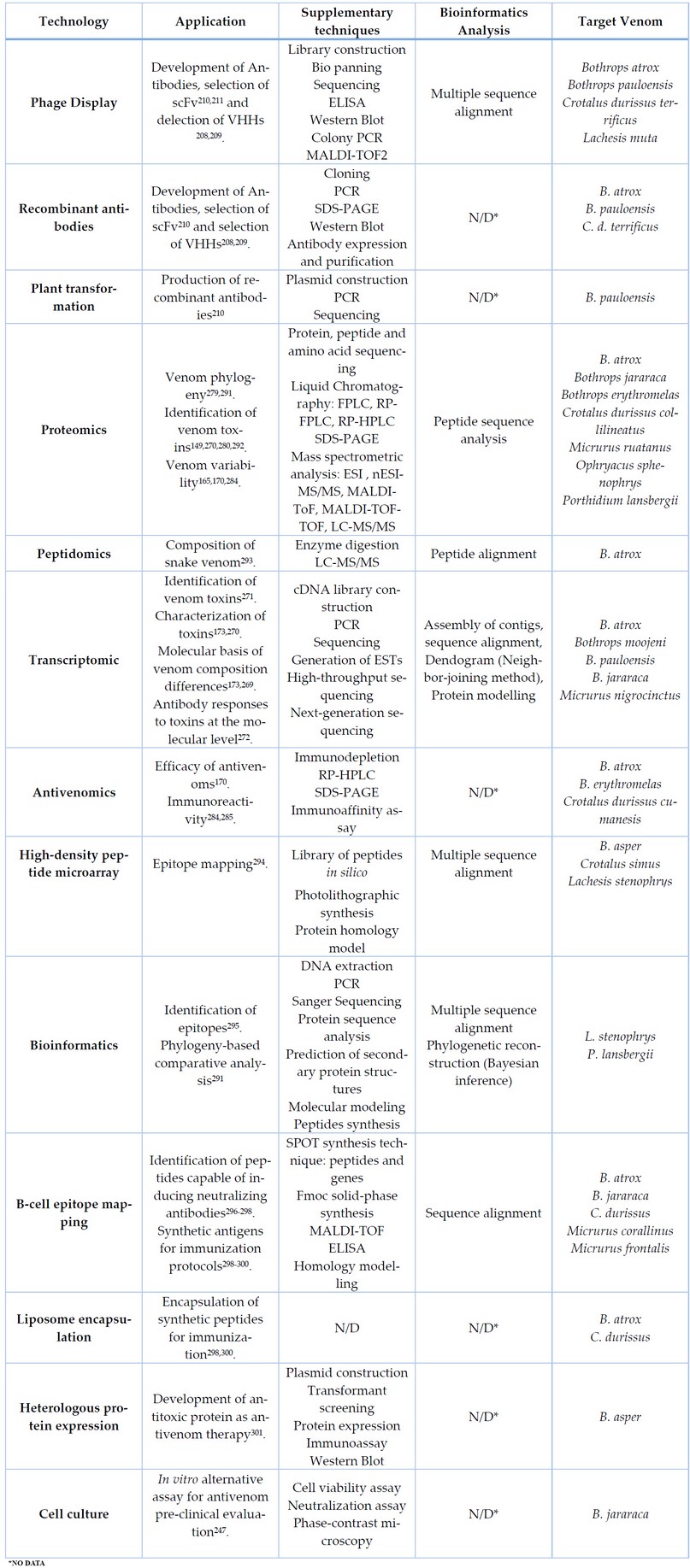
Table 4. Current biotechnological technologies for producing snake antivenoms in the Neotropics use bioinformatics tools.
Scientific evidence for decision-making in Public Health
The vast amount of information found in the published scientific literature on snakebite epidemiology, strategies and other associated issues can create confusion among decision-makers in governmental entities in the region and hinder the formulation of public health policies. Based on this systematic review, we propose actions that governments should implement according to their country's needs.
Start with creating a single national registry system to obtain a database on the epidemiology of snake bites. The information obtained will make it possible to know which groups are most affected. With this base information, strategic programs for prevention, control, monitoring, planning and research can be developed.
Follow-up programs for victims of venomous snakebites can reduce long-term sequelae. Prevention strategies should include educational programs and the provision of protective equipment in rural areas whose main activity is agriculture. Efforts should be made to strengthen the medical room for rapid action in snakebite emergencies.
Last but not least, governments should provide resources for the characterization of clinically meaningful snake venoms and promote research to create effective and low-cost diagnostic and therapeutic tools.
CONCLUSIONS
Snakebite disease is considered a neglected tropical disease due to its global, regional and national burden, as well as its social and economic impact on society. Although there are several prevention strategies and tools for treating this disease, a One Health approach is required because several actors are involved in its dynamics. Ecological, political, technological and medical aspects should be considered to allow us to manage and administer the correct registration of snakebite cases from public policies. It is essential to ensure the development of preventive programs and effective treatments for snakebite envenomation using current biotechnological tools for vulnerable populations. Preventive programs will improve the economic and social situation of the most affected regions today. The most modern biotechnological tools have been applied experimentally, but only on a laboratory scale, and the support of governmental entities is a crucial factor in enhancing the future industrial production and snakebite antivenoms scaled up.
Funding
Authors declare no such funding related to this article.
Acknowledgment
Not applicable.
Conflicts of Interest
Authors do not claim any conflict of interest.
REFERENCES
1. Kasturiratne A, Wickremasinghe AR, de Silva N, Gunawardena NK, Pathmeswaran A, Premaratna R, et al. The global burden of snakebite: a literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008;5(11):e218. doi:10.1371/journal.pmed.0050218
2. WHO. Snakebite Envenoming: A Strategy for Prevention and Control [Internet]. 2019 [cited 10 Jan 2022]. Available from: https://apps.who.int/iris/bitstream/handle/10665/324838/9789241515641-eng.pdf?ua=1
3. Jayawardana S, Gnanathasan A, Arambepola C, Chang T. Chronic Musculoskeletal Disabilities following Snake Envenoming in Sri Lanka: A Population-Based Study. PLoS Negl Trop Dis. 2016;10(11):e0005103. doi:10.1371/journal.pntd.0005103
4. Ebrahimi V, Hamdami E, Khademian MH, Moemenbellah-Fard MD, Vazirianzadeh B. Epidemiologic prediction of snake bites in tropical south Iran: Using seasonal time series methods. Clin Epidemiol Glob Health. 2018;6(4):208-15. doi:10.1016/j.cegh.2018.06.005
5. Chippaux JP. Incidence and mortality due to snakebite in the Americas. PLoS Negl Trop Dis. 2017;11(6):e0005662. doi:10.1371/journal.pntd.0005662
6. Engels D, Zhou XN. Neglected tropical diseases: an effective global response to local poverty-related disease priorities. Infect Dis Poverty. 2020;9(1):10. doi:10.1186/s40249-020-0630-9
7. Mechán Méndez V, Ramírez Erazo J, Wong Gutiérrez G, Durán Espinoza C, Meza Berrospi L, Mechán Escalante D, et al. Ceguera bilateral y manifestaciones hemato-neurológicas producidas durante un accidente botrópico. Acta Med Peruana. 2010;27(3):188-92.
8. Bawaskar HS, Bawaskar PH, Bawaskar PH. The global burden of snake bite envenoming. J R Coll Physicians Edinb. 2021;51(1):7-8. doi:10.4997/jrcpe.2021.102
9. Chippaux JP. [Global incidence of snake 0and scorpion envenoming]. Med Sci. 2009;25(2):197-200. doi:10.1051/medsci/2009252197
10. Gutiérrez JM. Improving antivenom availability and accessibility: science, technology, and beyond. Toxicon. 2012;60(4):676-87. doi:10.1016/j.toxicon.2012.02.008
11. Gutiérrez JM. Global Availability of Antivenoms: The Relevance of Public Manufacturing Laboratories. Toxins (Basel). 2018;11(1). doi:10.3390/toxins11010005
12. Longbottom J, Shearer FM, Devine M, Alcoba G, Chappuis F, Weiss DJ, et al. Vulnerability to snakebite envenoming: a global mapping of hotspots. Lancet. 2018;392(10148):673-84. doi:10.1016/s0140-6736(18)31224-8
13. Ortiz-Prado E, Yeager J, Andrade F, Schiavi-Guzman C, Abedrabbo-Figueroa P, Terán E, et al. Snake antivenom production in Ecuador: Poor implementation, and an unplanned cessation leads to a call for a renaissance. Toxicon. 2021;202:90-7. doi:10.1016/j.toxicon.2021.09.014
14. Williams HF, Layfield HJ, Vallance T, Patel K, Bicknell AB, Trim SA, et al. The Urgent Need to Develop Novel Strategies for the Diagnosis and Treatment of Snakebites. Toxins (Basel). 2019;11(6). doi:10.3390/toxins11060363
15. Campbell JA, Lamar WW, Brodie ED. The venomous reptiles of the western hemisphere. 1 ed. Comstock: Cornell University Press; 2004.
16. Warrell D. Snakebites in Central and South America: epidemiology, clinical features and clinical management. In: Campbell J, Lamar W, editors. The venomous reptiles of the Western hemisphere. Ithaca: Cornell University Press; 2004. p. 709-21.
17. Zacarias D, Loyola R. Climate change impacts on the distribution of venomous snakes and snakebite risk in Mozambique. Climatic Change. 2019;152(1):195-207. doi:10.1007/s10584-018-2338-4
18. Goldstein E, Erinjery JJ, Martin G, Kasturiratne A, Ediriweera DS, de Silva HJ, et al. Integrating human behavior and snake ecology with agent-based models to predict snakebite in high risk landscapes. PLoS Negl Trop Dis. 2021;15(1):e0009047. doi:10.1371/journal.pntd.0009047
19. de Silva HJ, Kasturiratne A, Pathmeswaran A, Lalloo DG. Snakebite: the true disease burden has yet to be determined. Ceylon Med J. 2013;58(3):93-5. doi:10.4038/cmj.v58i3.6101
20. Gutiérrez JM, Calvete JJ, Habib AG, Harrison RA, Williams DJ, Warrell DA. Snakebite envenoming. Nat Rev Dis Primers. 2017;3:17063. doi:10.1038/nrdp.2017.63
21. Pach S, Le Geyt J, Gutiérrez JM, Williams D, Maduwage KP, Habib AG, et al. Paediatric snakebite envenoming: the world's most neglected 'Neglected Tropical Disease'? Arch Dis Child. 2020;105(12):1135-9. doi:10.1136/archdischild-2020-319417
22. Yañez-Arenas C, Díaz-Gamboa L, Patrón-Rivero C, López-Reyes K, Chiappa-Carrara X. Estimating geographic patterns of ophidism risk in Ecuador. Neotrop Biodivers. 2018;4(1):55-61. doi:10.1080/23766808.2018.1454762
23. Gutiérrez JM, Burnouf T, Harrison RA, Calvete JJ, Brown N, Jensen SD, et al. A Call for Incorporating Social Research in the Global Struggle against Snakebite. PLoS Negl Trop Dis. 2015;9(9):e0003960. doi:10.1371/journal.pntd.0003960
24. Gutiérrez JM. Reducing the impact of snakebite envenoming in Latin America and the Caribbean: achievements and challenges ahead. Trans R Soc Trop Med Hyg. 2014;108(9):530-7. doi:10.1093/trstmh/tru102
25. Walter FG, Stolz U, Shirazi F, McNally J. Temporal analyses of coral snakebite severity published in the American Association of Poison Control Centers' Annual Reports from 1983 through 2007. Clin Toxicol. 2010;48(1):72-8. doi:10.3109/15563650903430944
26. Ochoa-Avilés A, Heredia-Andino OS, Escandón SA, Celorio-Carvajal CA, Arias-Peláez MC, Zaruma-Torres F, et al. Viperidae snakebites in Ecuador: A review of epidemiological and ecological aspects. Toxicon X. 2020;7:100051. doi:10.1016/j.toxcx.2020.100051
27. Chippaux JP, Williams V, White J. Snake venom variability: methods of study, results and interpretation. Toxicon. 1991;29(11):1279-303. doi:10.1016/0041-0101(91)90116-9
28. Shashidharamurthy R, Kemparaju K. Region-specific neutralization of Indian cobra (Naja naja) venom by polyclonal antibody raised against the eastern regional venom: A comparative study of the venoms from three different geographical distributions. Int Immunopharmacol. 2007;7(1):61-9. doi:10.1016/j.intimp.2006.08.014
29. Katibi OS, Adepoju FG, Olorunsola BO, Ernest SK, Monsudi KF. Blindness and scalp haematoma in a child following a snakebite. Afr Health Sci. 2015;15(3):1041-4. doi:10.4314/ahs.v15i3.46
30. Mustapha SK, Mubi BM, Askira BH. Bilateral blindness following snakebite. Trop Doct. 2010;40(2):117-8. doi:10.1258/td.2009.090429
31. Mao YC, Liu PY, Chiang LC, Lee CH, Lai CS, Lai KL, et al. Clinical manifestations and treatments of Protobothrops mucrosquamatus bite and associated factors for wound necrosis and subsequent debridement and finger or toe amputation surgery. Clin Toxicol. 2021;59(1):28-37. doi:10.1080/15563650.2020.1762892
32. Penington A, Johnstone B. A case of local tissue necrosis following a bite by the Australian tiger snake Notechis scutatus. Aust N Z J Surg. 1997;67(6):385-8. doi:10.1111/j.1445-2197.1997.tb01996.x
33. Del Brutto OH, Del Brutto VJ. Neurological complications of venomous snake bites: a review. Acta Neurol Scand. 2012;125(6):363-72. doi:10.1111/j.1600-0404.2011.01593.x
34. Gnanathasan A, Rodrigo C. Pulmonary effects and complications of snakebites. Chest. 2014;146(5):1403-12. doi:10.1378/chest.13-2674
35. Moseley T. Coral snake bite: recovery following symptoms of respiratory paralysis. Ann Surg. 1966;163(6):943-8. doi:10.1097/00000658-196606000-00016
36. Pinho FM, Zanetta DM, Burdmann EA. Acute renal failure after Crotalus durissus snakebite: a prospective survey on 100 patients. Kidney Int. 2005;67(2):659-67. doi:10.1111/j.1523-1755.2005.67122.x
37. Vikrant S, Jaryal A, Parashar A. Clinicopathological spectrum of snake bite-induced acute kidney injury from India. World J Nephrol. 2017;6(3):150-61. doi:10.5527/wjn.v6.i3.150
38. Chara K, Baccouche N, Turki O, Regaig K, Chaari A, Bahloul M, et al. A rare complication of viper envenomation: cardiac failure. A case report. Med Sante Trop. 2017;27(1):52-5. doi:10.1684/mst.2016.0636
39. Bon C. Serum therapy was discovered 100 years ago. Envenomings and Their Treatments: Fondation Marcel Mérieux; 1996. p. 3-9.
40. Knudsen C, Jürgensen JA, Føns S, Haack AM, Friis RUW, Dam SH, et al. Snakebite Envenoming Diagnosis and Diagnostics. Front Immunol. 2021;12:661457. doi:10.3389/fimmu.2021.661457
41. Ramos HR, Ho PL. Developing Snake Antivenom Sera by Genetic Immunization: A Review. In: Gopalakrishnakone P, Faiz A, Fernando R, Gnanathasan CA, Habib AG, Yang C-C, editors. Clinical Toxinology in Asia Pacific and Africa. Dordrecht: Springer Netherlands; 2015. p. 401-14.
42. Murphy JC. Antivenoms administered properly and swiftly save lives. Nat Med. 2010;16(7):747-. doi:10.1038/nm0710-747
43. WHO. WHO Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins [Internet]. 2016 [cited 18 Jan 2022]. Available from: https://www.who.int/biologicals/expert_committee/Antivenom_WHO_Guidelines_DJW_DEB_mn_cp.pdf
44. Alirahimi E, Kazemi-Lomedasht F, Shahbazzadeh D, Habibi-Anbouhi M, Hosseininejad Chafi M, Sotoudeh N, et al. Nanobodies as novel therapeutic agents in envenomation. Biochim Biophys Acta Gen Subj. 2018;1862(12):2955-65. doi:10.1016/j.bbagen.2018.08.019
45. Fernandes CFC, Pereira SS, Luiz MB, Silva N, Silva MCS, Marinho ACM, et al. Engineering of single-domain antibodies for next-generation snakebite antivenoms. Int J Biol Macromol. 2021;185:240-50. doi:10.1016/j.ijbiomac.2021.06.043
46. Srinivasa V, Sundaram MS, Anusha S, Hemshekhar M, Chandra Nayaka S, Kemparaju K, et al. Novel apigenin based small molecule that targets snake venom metalloproteases. PLoS One. 2014;9(9):e106364. doi:10.1371/journal.pone.0106364
47. Xie C, Slagboom J, Albulescu LO, Somsen GW, Vonk FJ, Casewell NR, et al. Neutralising effects of small molecule toxin inhibitors on nanofractionated coagulopathic Crotalinae snake venoms. Acta Pharm Sin B. 2020;10(10):1835-45. doi:10.1016/j.apsb.2020.09.005
48. Bermúdez-Méndez E, Fuglsang-Madsen A, Føns S, Lomonte B, Gutiérrez JM, Laustsen AH. Innovative Immunization Strategies for Antivenom Development. Toxins (Basel). 2018;10(11). doi:10.3390/toxins10110452
49. Harrison RA, Moura-Da-Silva AM, Laing GD, Wu Y, Richards A, Broadhead A, et al. Antibody from mice immunized with DNA encoding the carboxyl-disintegrin and cysteine-rich domain (JD9) of the haemorrhagic metalloprotease, Jararhagin, inhibits the main lethal component of viper venom. Clin Exp Immunol. 2000;121(2):358-63. doi:10.1046/j.1365-2249.2000.01287.x
50. Finney ER, Padula AM, Leister EM. Red-bellied black snake (Pseudechis porphyriacus) envenomation in 17 dogs: clinical signs, coagulation changes, haematological abnormalities, venom antigen levels and outcomes following treatment with a tiger-brown snake antivenom. Aust Vet J. 2020;98(7):319-25. doi:10.1111/avj.12953
51. Padula AM, Winkel KD. Red-bellied black snake (Pseudechis porphyriacus) envenomation in the dog: Diagnosis and treatment of nine cases. Toxicon. 2016;117:69-75. doi:10.1016/j.toxicon.2016.03.022
52. Tangtrongchitr T, Thumtecho S, Janprasert J, Sanprasert K, Tongpoo A, Tanpudsa Y, et al. Malayan Pit Viper Envenomation and Treatment in Thailand. Ther Clin Risk Manag. 2021;17:1257-66. doi:10.2147/tcrm.S337199
53. Bagnall L, Taliaferro D. Venomous Snakebites: Treatment Considerations and the Emergency Department Nurse Practitioners'/Providers' Priorities of Care. Adv Emerg Nurs J. 2021;43(4):309-23. doi:10.1097/tme.0000000000000374
54. Curran-Sills G, Kroeker J. Venomous Snakebites in Canada: A National Review of Patient Epidemiology and Antivenom Usage. Wilderness Environ Med. 2018;29(4):437-45. doi:10.1016/j.wem.2018.06.005
55. Gutiérrez JM, Fan HW. Improving the control of snakebite envenomation in Latin America and the Caribbean: a discussion on pending issues. Trans R Soc Trop Med Hyg. 2018;112(12):523-6. doi:10.1093/trstmh/try104
56. Habib AG, Musa BM, Iliyasu G, Hamza M, Kuznik A, Chippaux JP. Challenges and prospects of snake antivenom supply in sub-Saharan Africa. PLoS Negl Trop Dis. 2020;14(8):e0008374. doi:10.1371/journal.pntd.0008374
57. Mender MM, Bolton F, Berry C, Young M. Antivenom: An immunotherapy for the treatment of snakebite envenoming in sub-Saharan Africa. Adv Protein Chem Struct Biol. 2022;129:435-77. doi:10.1016/bs.apcsb.2021.11.004
58. Lamb T, de Haro L, Lonati D, Brvar M, Eddleston M. Antivenom for European Vipera species envenoming. Clin Toxicol (Phila). 2017;55(6):557-68. doi:10.1080/15563650.2017.1300261
59. Cai B, Huang Y, Chen YY, Hu JH, Guo XG, Wang YZ. Geographic patterns and ecological factors correlates of snake species richness in China. Dongwuxue Yanjiu. 2012;33(4):343-53. doi:10.3724/sp.J.1141.2012.04343
60. Dutto M, Lonati D, Goyffon M. Evidence of snakes in urban areas. Analysis of cases in the province of Cuneo and public health issues. Ann Ig. 2012;24(3):217-20.
61. Jasper EH, Miller M, Neuburger KJ, Widder PC, Snyder JW, Lopez BL. Venomous snakebites in an urban area: what are the possibilities? Wilderness Environ Med. 2000;11(3):168-71. doi:10.1580/1080-6032(2000)011[0168:vsiaua]2.3.co;2
62. Praba-Egge AD, Cone SW, Araim O, Freire I, Paida G, Escalante J, et al. Snakebites in the rainforests of Ecuador. World J Surg. 2003;27(2):234-40. doi:10.1007/s00268-002-6552-9
63. Halliday WD, Blouin-Demers G. Differential fitness in field and forest explains density-independent habitat selection by gartersnakes. Oecologia. 2016;181(3):841-51. doi:10.1007/s00442-016-3605-6
64. Silva JLD, Fonseca WLD, Mota da Silva A, Amaral G, Ortega GP, Oliveira AS, et al. Venomous snakes and people in a floodplain forest in the Western Brazilian Amazon: Potential risks for snakebites. Toxicon. 2020;187:232-44. doi:10.1016/j.toxicon.2020.09.007
65. Jones T, Babb R, Hensley F, LiWanPo C, Sullivan B. Sonoran desert snake communities at two sites: Concordance and effects of increased road traffic. Herpetol Conserv Biol. 2011;6:61-71.
66. Mendelson JR, Jennings WB. Shifts in the Relative Abundance of Snakes in a Desert Grassland. J Herpetol. 1992;26(1):38-45. doi:10.2307/1565019
67. Currie B. Urban envenomation. Med J Aust. 1989;151(11-12):724. doi:10.5694/j.1326-5377.1989.tb139678.x
68. Hosaka T, Numata S. Spatiotemporal dynamics of urban green spaces and human-wildlife conflicts in Tokyo. Sci Rep. 2016;6:30911. doi:10.1038/srep30911
69. Guedes TB, Sawaya RJ, Zizka A, Laffan S, Faurby S, Pyron RA, et al. Patterns, biases and prospects in the distribution and diversity of Neotropical snakes. Glob Ecol Biogeogr. 2018;27(1):14-21. doi:10.1111/geb.12679
70. Warrell DA. Venomous bites and stings in the tropical world. Med J Aust. 1993;159(11-12):773-9. doi:10.5694/j.1326-5377.1993.tb141345.x
71. Andrade-Díaz MS, Sarquis JA, Loiselle BA, Giraudo AR, Díaz-Gómez JM. Expansion of the agricultural frontier in the largest South American Dry Forest: Identifying priority conservation areas for snakes before everything is lost. PLoS One. 2019;14(9):e0221901. doi:10.1371/journal.pone.0221901
72. Vigne JD. The origins of animal domestication and husbandry: a major change in the history of humanity and the biosphere. C R Biol. 2011;334(3):171-81. doi:10.1016/j.crvi.2010.12.009
73. Wells JCK, Stock JT. Life History Transitions at the Origins of Agriculture: A Model for Understanding How Niche Construction Impacts Human Growth, Demography and Health. Front Endocrinol. 2020;11:325. doi:10.3389/fendo.2020.00325
74. Zeder MA. Domestication and early agriculture in the Mediterranean Basin: Origins, diffusion, and impact. Proc Natl Acad Sci U S A. 2008;105(33):11597-604. doi:10.1073/pnas.0801317105
75. Jayawardana S, Arambepola C, Chang T, Gnanathasan A. Prevalence, vulnerability and epidemiological characteristics of snakebite in agricultural settings in rural Sri Lanka: A population-based study from South Asia. PLoS One. 2020;15(12):e0243991. doi:10.1371/journal.pone.0243991
76. Mise YF, Lira-da-Silva RM, Carvalho FM. Agriculture and snakebite in Bahia, Brazil - An ecological study. Ann Agric Environ Med. 2016;23(3):416-9. doi:10.5604/12321966.1219179
77. Mise YF, Lira-da-Silva RM, Carvalho FM. Fatal Snakebite Envenoming and Agricultural Work in Brazil: A Case-Control Study. Am J Trop Med Hyg. 2019;100(1):150-4. doi:10.4269/ajtmh.18-0579
78. Suazo-Ortuño I, Alvarado-Díaz J, Martínez-Ramos M. Effects of conversion of dry tropical forest to agricultural mosaic on herpetofaunal assemblages. Conserv Biol. 2008;22(2):362-74. doi:10.1111/j.1523-1739.2008.00883.x
79. Ochoa C, Bolon I, Durso AM, Ruiz de Castañeda R, Alcoba G, Babo Martins S, et al. Assessing the Increase of Snakebite Incidence in Relationship to Flooding Events. J Environ Public Health. 2020;2020:6135149. doi:10.1155/2020/6135149
80. 80. González-Andrade F, Chippaux JP. Snake bite envenomation in Ecuador. Trans R Soc Trop Med Hyg. 2010;104(9):588-91. doi:10.1016/j.trstmh.2010.05.006
81. Needleman RK, Neylan IP, Erickson T. Potential Environmental and Ecological Effects of Global Climate Change on Venomous Terrestrial Species in the Wilderness. Wilderness Environ Med. 2018;29(2):226-38. doi:10.1016/j.wem.2017.11.004
82. Lourenço-de-Moraes R, Lansac-Toha FM, Schwind LTF, Arrieira RL, Rosa RR, Terribile LC, et al. Climate change will decrease the range size of snake species under negligible protection in the Brazilian Atlantic Forest hotspot. Sci Rep. 2019;9(1):8523. doi:10.1038/s41598-019-44732-z
83. Martín G, Yáñez-Arenas C, Rangel-Camacho R, Murray KA, Goldstein E, Iwamura T, et al. Implications of global environmental change for the burden of snakebite. Toxicon X. 2021;9-10:100069. doi:10.1016/j.toxcx.2021.100069
84. Martinez PA, Andrade MA, Bidau CJ. Potential effects of climate change on the risk of accidents with poisonous species of the genus Tityus (Scorpiones, Buthidae) in Argentina. Spat Spatiotemporal Epidemiol. 2018;25:67-72. doi:10.1016/j.sste.2018.03.002
85. Delgado ABT, Gondim C, Reichert LP, da Silva PHV, Souza R, Fernandes TMP, et al. Hemorrhagic stroke secondary to Bothrops spp. venom: A case report. Toxicon. 2017;132:6-8. doi:10.1016/j.toxicon.2017.03.015
86. Yoshida-Kanashiro E, Navarrete LF, Rodríguez-Acosta A. On the unsual hemorrhagic and necrotic activities caused by the rattlesnake (Crotalus durissus cumanensis) in a Venezuelan patient. Rev Cubana Med Trop. 2003;55(1):38-40.
87. Ribeiro LA, Jorge MT, Lebrão ML. Prognostic factors for local necrosis in Bothrops jararaca (Brazilian pit viper) bites. Trans R Soc Trop Med Hyg. 2001;95(6):630-4. doi:10.1016/s0035-9203(01)90101-4
88. Saborío P, González M, Cambronero M. [Snake bite accidents in children in Costa Rica: epidemiology and determination of risk factors in the development of abscess and necrosis]. Toxicon. 1998;36(2):359-66. doi:10.1016/s0041-0101(97)00076-7
89. Albuquerque PL, Silva GB, Jr., Jacinto CN, Lima JB, Lima CB, Amaral YS, et al. Acute kidney injury after snakebite accident treated in a Brazilian tertiary care centre. Nephrology. 2014;19(12):764-70. doi:10.1111/nep.12327
90. Torrez PP, Said R, Quiroga MM, Duarte MR, França FO. Forest pit viper (Bothriopsis bilineata bilineata) bite in the Brazilian Amazon with acute kidney injury and persistent thrombocytopenia. Toxicon. 2014;85:27-30. doi:10.1016/j.toxicon.2014.04.001
91. Bhaumik S, Kallakuri S, Kaur A, Devarapalli S, Daniel M. Mental health conditions after snakebite: a scoping review. BMJ Glob Health. 2020;5(11). doi:10.1136/bmjgh-2020-004131
92. Wijesinghe CA, Williams SS, Kasturiratne A, Dolawaththa N, Wimalaratne P, Wijewickrema B, et al. A Randomized Controlled Trial of a Brief Intervention for Delayed Psychological Effects in Snakebite Victims. PLoS Negl Trop Dis. 2015;9(8):e0003989. doi:10.1371/journal.pntd.0003989
93. Silva KV, Said RDC, Assy J, Duarte MR, Torrez PPQ, França FOS. A case of envenomation caused by Oxybelis fulgidus (Serpentes, Colubridae) in Brazilian Amazon. Rev Soc Bras Med Trop. 2019;52:e20180423. doi:10.1590/0037-8682-0426-2018
94. Liapis K, Charitaki E, Psaroulaki A. Case Report: Spherocytic Hemolytic Anemia after Envenomation by Long-Nosed Viper (Vipera ammodytes). Am J Trop Med Hyg. 2019;101(6):1442-5. doi:10.4269/ajtmh.19-0611
95. Resiere D, Houcke S, Pujo JM, Mayence C, Mathien C, NkontCho F, et al. Clinical Features and Management of Snakebite Envenoming in French Guiana. Toxins (Basel). 2020;12(10). doi:10.3390/toxins12100662
96. Brenes-Chacón H, Ulloa-Gutierrez R, Soriano-Fallas A, Camacho-Badilla K, Valverde-Muñoz K, Ávila-Agüero ML. Bacterial Infections Associated with Viperidae Snakebites in Children: A 14-Year Experience at the Hospital Nacional de Niños de Costa Rica(†). Am J Trop Med Hyg. 2019;100(5):1227-9. doi:10.4269/ajtmh.18-1015
97. Liu PY, Shi ZY, Lin CF, Huang JA, Liu JW, Chan KW, et al. Shewanella infection of snake bites: a twelve-year retrospective study. Clinics. 2012;67(5):431-5. doi:10.6061/clinics/2012(05)05
98. Mamede CCN, de Sousa Simamoto BB, da Cunha Pereira DF, de Oliveira Costa J, Ribeiro MSM, de Oliveira F. Edema, hyperalgesia and myonecrosis induced by Brazilian bothropic venoms: overview of the last decade. Toxicon. 2020;187:10-8. doi:10.1016/j.toxicon.2020.08.016
99. Jayawardana S, Arambepola C, Chang T, Gnanathasan A. Long-term health complications following snake envenoming. J Multidiscip Healthc. 2018;11:279-85. doi:10.2147/jmdh.S126648
100. Brenes-Chacon H, Gutierrez JM, Camacho-Badilla K, Soriano-Fallas A, Ulloa-Gutierrez R, Valverde K, et al. Long-term sequelae secondary to snakebite envenoming: a single centre retrospective study in a Costa Rican paediatric hospital. BMJ Paediatr Open. 2020;4(1):e000735. doi:10.1136/bmjpo-2020-000735
101. Dhar D. Compartment Syndrome Following Snake Bite. Oman Med J. 2015;30(2):e082. doi:10.5001/omj.2015.32
102. Abubakar SB, Habib AG, Mathew J. Amputation and disability following snakebite in Nigeria. Trop Doct. 2010;40(2):114-6. doi:10.1258/td.2009.090266
103. George TK, Toms AG, Fenn BN, Kumar V, Kavitha R, Georgy JT, et al. Renal outcomes among snake-envenomed patients with acute kidney injury in southern India. Natl Med J India. 2019;32(1):5-8. doi:10.4103/0970-258x.272106
104. Naqvi R. Snake-bite-induced Acute Kidney Injury. J Coll Physicians Surg Pak. 2016;26(6):517-20.
105. Sarkar S, Sinha R, Chaudhury AR, Maduwage K, Abeyagunawardena A, Bose N, et al. Snake bite associated with acute kidney injury. Pediatr Nephrol. 2020;36(12):3829-40. doi:10.1007/s00467-020-04911-x
106. Bogarín G, Morais JF, Yamaguchi IK, Stephano MA, Marcelino JR, Nishikawa AK, et al. Neutralization of crotaline snake venoms from Central and South America by antivenoms produced in Brazil and Costa Rica. Toxicon. 2000;38(10):1429-41. doi:10.1016/s0041-0101(99)00236-6
107. Habib ZG, Salihu AS, Hamza M, Yakasai AM, Iliyasu G, Yola IM, et al. Posttraumatic stress disorder and psycho-social impairment following snakebite in Northeastern Nigeria. Int J Psychiatry Med. 2021;56(2):97-115. doi:10.1177/0091217420913400
108. Muhammed A, Dalhat MM, Joseph BO, Ahmed A, Nguku P, Poggensee G, et al. Predictors of depression among patients receiving treatment for snakebite in General Hospital, Kaltungo, Gombe State, Nigeria: August 2015. Int J Ment Health Syst. 2017;11:26. doi:10.1186/s13033-017-0132-8
109. Williams SS, Wijesinghe CA, Jayamanne SF, Buckley NA, Dawson AH, Lalloo DG, et al. Delayed psychological morbidity associated with snakebite envenoming. PLoS Negl Trop Dis. 2011;5(8):e1255. doi:10.1371/journal.pntd.0001255
110. Khosrojerdi H, Amini M. Acute and Delayed Stress Symptoms Following Snakebite. Asia Pac J Med Toxicol. 2013;2(4):140-4. doi:10.22038/apjmt.2013.2037
111. Harrison RA, Hargreaves A, Wagstaff SC, Faragher B, Lalloo DG. Snake envenoming: a disease of poverty. PLoS Negl Trop Dis. 2009;3(12):e569. doi:10.1371/journal.pntd.0000569
112. Arias-Rodríguez J, Gutiérrez JM. Circumstances and Consequences of Snakebite Envenomings: A Qualitative Study in South-Eastern Costa Rica. Toxins (Basel). 2020;12(1). doi:10.3390/toxins12010045
113. Hasan SM, Basher A, Molla AA, Sultana NK, Faiz MA. The impact of snake bite on household economy in Bangladesh. Trop Doct. 2012;42(1):41-3. doi:10.1258/td.2011.110137
114. Vaiyapuri S, Vaiyapuri R, Ashokan R, Ramasamy K, Nattamaisundar K, Jeyaraj A, et al. Snakebite and its socio-economic impact on the rural population of Tamil Nadu, India. PLoS One. 2013;8(11):e80090. doi:10.1371/journal.pone.0080090
115. Habib AG, Brown NI. The snakebite problem and antivenom crisis from a health-economic perspective. Toxicon. 2018;150:115-23. doi:10.1016/j.toxicon.2018.05.009
116. Pinho FM, Burdmann EA. Fatal cerebral hemorrhage and acute renal failure after young Bothrops jararacussu snake bite. Ren Fail. 2001;23(2):269-77. doi:10.1081/jdi-100103499
117. Lizarazo J, Patiño R, Lizarazo D, Osorio G. Fatal brain hemorrhage after Bothrops asper bite in the Catatumbo region of Colombia. Biomedica. 2020;40(4):609-15. doi:10.7705/biomedica.5181
118. Gutiérrez JM. Envenenamientos por mordeduras de serpientes en América Latina y el Caribe: Una visión integral de carácter regional. J Boletín de Malariología y Salud Ambiental. 2011;51:1-16.
119. Swaroop S, Grab B. Snakebite mortality in the world. Bull World Health Organ. 1954;10(1):35-76.
120. Chippaux JP. Snake-bites: appraisal of the global situation. Bull World Health Organ. 1998;76(5):515-24.
121. Gutiérrez JM, Williams D, Fan HW, Warrell DA. Snakebite envenoming from a global perspective: Towards an integrated approach. Toxicon. 2010;56(7):1223-35. doi:10.1016/j.toxicon.2009.11.020
122. Habib AG, Kuznik A, Hamza M, Abdullahi MI, Chedi BA, Chippaux JP, et al. Snakebite is Under Appreciated: Appraisal of Burden from West Africa. PLoS Negl Trop Dis. 2015;9(9):e0004088. doi:10.1371/journal.pntd.0004088
123. Gutiérrez JM, Maduwage K, Iliyasu G, Habib A. Snakebite envenoming in different national contexts: Costa Rica, Sri Lanka, and Nigeria. Toxicon X. 2021;9-10:100066. doi:10.1016/j.toxcx.2021.100066
124. Giovannini P. Medicinal plants of the Achuar (Jivaro) of Amazonian Ecuador: ethnobotanical survey and comparison with other Amazonian pharmacopoeias. J Ethnopharmacol. 2015;164:78-88. doi:10.1016/j.jep.2015.01.038
125. Giovannini P, Howes MR. Medicinal plants used to treat snakebite in Central America: Review and assessment of scientific evidence. J Ethnopharmacol. 2017;199:240-56. doi:10.1016/j.jep.2017.02.011
126. Chaves LF, Chuang TW, Sasa M, Gutiérrez JM. Snakebites are associated with poverty, weather fluctuations, and El Niño. Sci Adv. 2015;1(8):e1500249. doi:10.1126/sciadv.1500249
127. Squaiella-Baptistão CC, Sant'Anna OA, Marcelino JR, Tambourgi DV. The history of antivenoms development: Beyond Calmette and Vital Brazil. Toxicon. 2018;150:86-95. doi:10.1016/j.toxicon.2018.05.008
128. Gutiérrez JM, Teixeira CFP, Fan HW. Instituto Butantan and Instituto Clodomiro Picado: A long-standing partnership in science, technology, and public health. Toxicon. 2021;202:75-81. doi:10.1016/j.toxicon.2021.09.007
129. Gutiérrez JM, Higashi HG, Wen FH, Burnouf T. Strengthening antivenom production in Central and South American public laboratories: report of a workshop. Toxicon. 2007;49(1):30-5. doi:10.1016/j.toxicon.2006.09.005
130. Gutiérrez JM, Lomonte B, Sanz L, Calvete JJ, Pla D. Immunological profile of antivenoms: preclinical analysis of the efficacy of a polyspecific antivenom through antivenomics and neutralization assays. J Proteomics. 2014;105:340-50. doi:10.1016/j.jprot.2014.02.021
131. Ainsworth S, Menzies SK, Casewell NR, Harrison RA. An analysis of preclinical efficacy testing of antivenoms for sub-Saharan Africa: Inadequate independent scrutiny and poor-quality reporting are barriers to improving snakebite treatment and management. PLoS Negl Trop Dis. 2020;14(8):e0008579. doi:10.1371/journal.pntd.0008579
132. Baldé MC, Chippaux JP, Boiro MY, Stock RP, Massougbodji A. Use of antivenoms for the treatment of envenomation by Elapidae snakes in Guinea, Sub-Saharan Africa. J Venom Anim Toxins Incl Trop Dis. 2013;19(1):6. doi:10.1186/1678-9199-19-6
133. Gutiérrez JM. Preclinical assessment of the neutralizing efficacy of snake antivenoms in Latin America and the Caribbean: A review. Toxicon. 2018;146:138-50. doi:10.1016/j.toxicon.2018.02.053
134. Zdenek CN, Hay C, Arbuckle K, Jackson TNW, Bos MHA, Op den Brouw B, et al. Coagulotoxic effects by brown snake (Pseudonaja) and taipan (Oxyuranus) venoms, and the efficacy of a new antivenom. Toxicol In Vitro. 2019;58:97-109. doi:10.1016/j.tiv.2019.03.031
135. Resiere D, Mehdaoui H, Gutiérrez JM. Snakebite envenomation in the Caribbean: The role of medical and scientific cooperation. PLoS Negl Trop Dis. 2018;12(7):e0006441. doi:10.1371/journal.pntd.0006441
136. Cardoso JL, Fan HW, França FO, Jorge MT, Leite RP, Nishioka SA, et al. Randomized comparative trial of three antivenoms in the treatment of envenoming by lance-headed vipers (Bothrops jararaca) in São Paulo, Brazil. Q J Med. 1993;86(5):315-25.
137. Otero-Patiño R, Segura A, Herrera M, Angulo Y, León G, Gutiérrez JM, et al. Comparative study of the efficacy and safety of two polyvalent, caprylic acid fractionated [IgG and F(ab')2] antivenoms, in Bothrops asper bites in Colombia. Toxicon. 2012;59(2):344-55. doi:10.1016/j.toxicon.2011.11.017
138. Pardal PP, Souza SM, Monteiro MR, Fan HW, Cardoso JL, França FO, et al. Clinical trial of two antivenoms for the treatment of Bothrops and Lachesis bites in the north eastern Amazon region of Brazil. Trans R Soc Trop Med Hyg. 2004;98(1):28-42. doi:10.1016/s0035-9203(03)00005-1
139. Resiere D, Villalta M, Arias AS, Kallel H, Nèviére R, Vidal N, et al. Snakebite envenoming in French Guiana: Assessment of the preclinical efficacy against the venom of Bothrops atrox of two polyspecific antivenoms. Toxicon. 2020;173:1-4. doi:10.1016/j.toxicon.2019.11.001
140. Madrigal M, Pla D, Sanz L, Barboza E, Arroyo-Portilla C, Corrêa-Netto C, et al. Cross-reactivity, antivenomics, and neutralization of toxic activities of Lachesis venoms by polyspecific and monospecific antivenoms. PLoS Negl Trop Dis. 2017;11(8):e0005793. doi:10.1371/journal.pntd.0005793
141. Segura A, Castillo MC, Núñez V, Yarlequé A, Gonçalves LR, Villalta M, et al. Preclinical assessment of the neutralizing capacity of antivenoms produced in six Latin American countries against medically-relevant Bothrops snake venoms. Toxicon. 2010;56(6):980-9. doi:10.1016/j.toxicon.2010.07.001
142. Smalligan R, Cole J, Brito N, Laing GD, Mertz BL, Manock S, et al. Crotaline snake bite in the Ecuadorian Amazon: randomised double blind comparative trial of three South American polyspecific antivenoms. Bmj. 2004;329(7475):1129. doi:10.1136/bmj.329.7475.1129
143. de Roodt AR, Lanari LC, de Oliveira VC, Laskowicz RD, Stock RP. Neutralization of Bothrops alternatus regional venom pools and individual venoms by antivenom: a systematic comparison. Toxicon. 2011;57(7-8):1073-80. doi:10.1016/j.toxicon.2011.04.013
144. Leiva CL, Cangelosi A, Mariconda V, Farace M, Geoghegan P, Brero L, et al. IgY-based antivenom against Bothrops alternatus: Production and neutralization efficacy. Toxicon. 2019;163:84-92. doi:10.1016/j.toxicon.2019.03.020
145. Sousa LF, Nicolau CA, Peixoto PS, Bernardoni JL, Oliveira SS, Portes-Junior JA, et al. Comparison of phylogeny, venom composition and neutralization by antivenom in diverse species of Bothrops complex. PLoS Negl Trop Dis. 2013;7(9):e2442. doi:10.1371/journal.pntd.0002442
146. Baudou FG, Litwin S, Lanari LC, Laskowicz RD, Damin CF, Chippaux JP, et al. Antivenom against Crotalus durissus terrificus venom: Immunochemical reactivity and experimental neutralizing capacity. Toxicon. 2017;140:11-7. doi:10.1016/j.toxicon.2017.10.009
147. Ponce-López R, Neri-Castro E, Borja M, Strickland JL, Alagón A. Neutralizing potency and immunochemical evaluation of an anti-Crotalus mictlantecuhtli experimental serum. Toxicon. 2020;187:171-80. doi:10.1016/j.toxicon.2020.08.026
148. Sánchez M, Solano G, Vargas M, Reta-Mares F, Neri-Castro É, Alagón A, et al. Toxicological profile of medically relevant Crotalus species from Mexico and their neutralization by a Crotalus basiliscus/Bothrops asper antivenom. Toxicon. 2020;179:92-100. doi:10.1016/j.toxicon.2020.03.006
149. Lippa E, Török F, Gómez A, Corrales G, Chacón D, Sasa M, et al. First look into the venom of Roatan Island's critically endangered coral snake Micrurus ruatanus: Proteomic characterization, toxicity, immunorecognition and neutralization by an antivenom. J Proteomics. 2019;198:177-85. doi:10.1016/j.jprot.2019.01.007
150. Rey-Suárez P, Lomonte B. Immunological cross-recognition and neutralization studies of Micrurus mipartitus and Micrurus dumerilii venoms by two therapeutic equine antivenoms. Biologicals. 2020;68:40-5. doi:10.1016/j.biologicals.2020.08.011
151. Rey-Suárez P, Núñez V, Fernández J, Lomonte B. Integrative characterization of the venom of the coral snake Micrurus dumerilii (Elapidae) from Colombia: Proteome, toxicity, and cross-neutralization by antivenom. J Proteomics. 2016;136:262-73. doi:10.1016/j.jprot.2016.02.006
152. Muniz EG, Noronha M, Saraiva M, Monteiro WM, Oliveira SS. Neutralization of hemostatic disorders induced by Lachesis muta venom using Brazilian antivenoms. Toxicon. 2021;191:44-7. doi:10.1016/j.toxicon.2020.12.013
153. FDA. Approved Blood Products, Coral Snake Antivenom [Internet]. 2018 [cited 15 Jan 2022]. Available from: https://www.fda.gov/vaccines-blood-biologics/approved-blood-products/coral-snake-antivenom
154. FDA. Approved Blood Products, CroFab [Internet]. 2018 [cited 15 Jan 2022]. Available from: https://www.fda.gov/vaccines-blood-biologics/approved-blood-products/crofab
155. FDA. Approved Blood Products, ANAVIP [Internet]. 2021 [cited 15 Jan 2022]. Available from: https://www.fda.gov/vaccines-blood-biologics/approved-blood-products/anavip
156. Martinez-Miranda E, Cabello Rosales A, Ortiz E. Valuation of Biotechnological Research: A Real Options Application for a Mexican Company. J Bus Econ Manag. 2018;10(2):1824-43.
157. FDA. Orphan Drug 2012 [cited Available from: https://www.fda.gov/media/83372/download#:~:text=Orphan%20Drug%20Definition%20An%20orphan,following%20approval%20by%20the%20FDA.
158. Waheed H, Moin SF, Choudhary MI. Snake Venom: From Deadly Toxins to Life-saving Therapeutics. Curr Med Chem. 2017;24(17):1874-91. doi:10.2174/0929867324666170605091546
159. Gómez-Betancur I, Gogineni V, Salazar-Ospina A, León F. Perspective on the Therapeutics of Anti-Snake Venom. Molecules. 2019;24(18):1-29. doi:10.3390/molecules24183276
160. Fan H, Vigilato M, Pompei J, JM G, RELAPA R. Situación de los laboratorios públicos productores de antivenenos en América Latina [Internet]. Panamerican Journal of Public Health; 2019 [cited 16 Dec 2021]. Available from: https://iris.paho.org/handle/10665.2/51693
161. Gutiérrez JM, Fan HW, Silvera CL, Angulo Y. Stability, distribution and use of antivenoms for snakebite envenomation in Latin America: report of a workshop. Toxicon. 2009;53(6):625-30. doi:10.1016/j.toxicon.2009.01.020
162. Gutiérrez JM, Zanette L, Vigilato MAN, Pompei JCA, Martins D, Fan HW. Appraisal of antivenom production in public laboratories in Latin America during the first semester of 2020: The impact of COVID-19. PLoS Negl Trop Dis. 2021;15(6):e0009469. doi:10.1371/journal.pntd.0009469
163. Kerrigan KR. Venomous snakebite in eastern Ecuador. Am J Trop Med Hyg. 1991;44(1):93-9. doi:10.4269/ajtmh.1991.44.93
164. MSP. Gaceta Epidemiológica Ecuador SIVE-ALERTA [Internet]. 2018 [cited 18 Nov 2021]. Available from: https://www.salud.gob.ec/gaceta-epidemiologica-ecuador-sive-alerta/
165. Oliveira IS, Cardoso IA, Bordon KCF, Carone SEI, Boldrini-França J, Pucca MB, et al. Global proteomic and functional analysis of Crotalus durissus collilineatus individual venom variation and its impact on envenoming. J Proteomics. 2019;191:153-65. doi:10.1016/j.jprot.2018.02.020
166. Larrick JW, Yost JA, Kaplan J. Snake bite among the Waorani Indians of Eastern Ecuador. Trans R Soc Trop Med Hyg. 1978;72(5):542-3. doi:10.1016/0035-9203(78)90184-0
167. Prior JE, Lynch MD, Gill RT. Broad-host-range vectors for protein expression across gram negative hosts. Biotechnol Bioeng. 2010;106(2):326-32. doi:10.1002/bit.22695
168. Theakston RD, Reid HA, Larrick JW, Kaplan J, Yost JA. Snake venom antibodies in Ecuadorian Indians. J Trop Med Hyg. 1981;84(5):199-202.
169. Heard K, O'Malley GF, Dart RC. Antivenom therapy in the Americas. Drugs. 1999;58(1):5-15. doi:10.2165/00003495-199958010-00002
170. Calvete JJ, Sanz L, Pérez A, Borges A, Vargas AM, Lomonte B, et al. Snake population venomics and antivenomics of Bothrops atrox: Paedomorphism along its transamazonian dispersal and implications of geographic venom variability on snakebite management. J Proteomics. 2011;74(4):510-27. doi:10.1016/j.jprot.2011.01.003
171. Vélez SM, Salazar M, Acosta de Patiño H, Gómez L, Rodriguez A, Correa D, et al. Geographical variability of the venoms of four populations of Bothrops asper from Panama: Toxicological analysis and neutralization by a polyvalent antivenom. Toxicon. 2017;132:55-61. doi:10.1016/j.toxicon.2017.04.002
172. Saldarriaga MM, Otero R, Núñez V, Toro MF, Díaz A, Gutiérrez JM. Ontogenetic variability of Bothrops atrox and Bothrops asper snake venoms from Colombia. Toxicon. 2003;42(4):405-11. doi:10.1016/s0041-0101(03)00171-5
173. Zelanis A, Andrade-Silva D, Rocha MM, Furtado MF, Serrano SM, Junqueira-de-Azevedo IL, et al. A transcriptomic view of the proteome variability of newborn and adult Bothrops jararaca snake venoms. PLoS Negl Trop Dis. 2012;6(3):e1554. doi:10.1371/journal.pntd.0001554
174. Munawar A, Ali SA, Akrem A, Betzel C. Snake Venom Peptides: Tools of Biodiscovery. Toxins (Basel). 2018;10(11). doi:10.3390/toxins10110474
175. Barua A, Mikheyev A. Many options, few solutions: over 60 million years snakes converged on a few optimal venom formulations. Molecular Biology and Evolution. 2019;36. doi:10.1093/molbev/msz125
176. Hiu JJ, Yap MKK. Cytotoxicity of snake venom enzymatic toxins: phospholipase A2 and l-amino acid oxidase. Biochem Soc Trans. 2020;48(2):719-31. doi:10.1042/bst20200110
177. Zhang C, Medzihradszky KF, Sánchez EE, Basbaum AI, Julius D. Lys49 myotoxin from the Brazilian lancehead pit viper elicits pain through regulated ATP release. Proc Natl Acad Sci U S A. 2017;114(12):E2524-e32. doi:10.1073/pnas.1615484114
178. Gutiérrez JM, Escalante T, Rucavado A, Herrera C, Fox JW. A Comprehensive View of the Structural and Functional Alterations of Extracellular Matrix by Snake Venom Metalloproteinases (SVMPs): Novel Perspectives on the Pathophysiology of Envenoming. Toxins (Basel). 2016;8(10). doi:10.3390/toxins8100304
179. Olaoba OT, Karina Dos Santos P, Selistre-de-Araujo HS, Ferreira de Souza DH. Snake Venom Metalloproteinases (SVMPs): A structure-function update. Toxicon X. 2020;7:100052. doi:10.1016/j.toxcx.2020.100052
180. Amorim FG, Menaldo DL, Carone SEI, Silva TA, Sartim MA, De Pauw E, et al. New Insights on Moojase, a Thrombin-Like Serine Protease from Bothrops moojeni Snake Venom. Toxins (Basel). 2018;10(12). doi:10.3390/toxins10120500
181. Boldrini-França J, Pinheiro-Junior EL, Peigneur S, Pucca MB, Cerni FA, Borges RJ, et al. Beyond hemostasis: a snake venom serine protease with potassium channel blocking and potential antitumor activities. Sci Rep. 2020;10(1):4476. doi:10.1038/s41598-020-61258-x
182. Nirthanan S. Snake three-finger α-neurotoxins and nicotinic acetylcholine receptors: molecules, mechanisms and medicine. Biochem Pharmacol. 2020;181:114168. doi:10.1016/j.bcp.2020.114168
183. Xie C, Bittenbinder MA, Slagboom J, Arrahman A, Bruijns S, Somsen GW, et al. Erythrocyte haemotoxicity profiling of snake venom toxins after nanofractionation. J Chromatogr B Analyt Technol Biomed Life Sci. 2021;1176:122586. doi:10.1016/j.jchromb.2021.122586
184. Guidolin FR, Caricati CP, Marcelino JR, da Silva WD. Development of Equine IgG Antivenoms against Major Snake Groups in Mozambique. PLoS Negl Trop Dis. 2016;10(1):e0004325. doi:10.1371/journal.pntd.0004325
185. Morais JF, de Freitas MC, Yamaguchi IK, dos Santos MC, da Silva WD. Snake antivenoms from hyperimmunized horses: comparison of the antivenom activity and biological properties of their whole IgG and F(ab')2 fragments. Toxicon. 1994;32(6):725-34. doi:10.1016/0041-0101(94)90341-7
186. Fernández GP, Segura A, Herrera M, Velasco W, Solano G, Gutiérrez JM, et al. Neutralization of Bothrops mattogrossensis snake venom from Bolivia: experimental evaluation of llama and donkey antivenoms produced by caprylic acid precipitation. Toxicon. 2010;55(2-3):642-5. doi:10.1016/j.toxicon.2009.07.031
187. Johnson PN, McGoodwin L, Banner W, Jr. Utilisation of Crotalidae polyvalent immune fab (ovine) for Viperidae envenomations in children. Emerg Med J. 2008;25(12):793-8. doi:10.1136/emj.2007.054916
188. Karlson-Stiber C, Persson H, Heath A, Smith D, al-Abdulla IH, Sjöström L. First clinical experiences with specific sheep Fab fragments in snake bite. Report of a multicentre study of Vipera berus envenoming. J Intern Med. 1997;241(1):53-8. doi:10.1046/j.1365-2796.1997.80896000.x
189. Laustsen AH, María Gutiérrez J, Knudsen C, Johansen KH, Bermúdez-Méndez E, Cerni FA, et al. Pros and cons of different therapeutic antibody formats for recombinant antivenom development. Toxicon. 2018;146:151-75. doi:10.1016/j.toxicon.2018.03.004
190. León G, Herrera M, Segura Á, Villalta M, Vargas M, Gutiérrez JM. Pathogenic mechanisms underlying adverse reactions induced by intravenous administration of snake antivenoms. Toxicon. 2013;76:63-76. doi:10.1016/j.toxicon.2013.09.010
191. Schaeffer TH, Khatri V, Reifler LM, Lavonas EJ. Incidence of immediate hypersensitivity reaction and serum sickness following administration of Crotalidae polyvalent immune Fab antivenom: a meta-analysis. Acad Emerg Med. 2012;19(2):121-31. doi:10.1111/j.1553-2712.2011.01276.x
192. de Silva HA, Ryan NM, de Silva HJ. Adverse reactions to snake antivenom, and their prevention and treatment. Br J Clin Pharmacol. 2016;81(3):446-52. doi:10.1111/bcp.12739
193. Malasit P, Warrell DA, Chanthavanich P, Viravan C, Mongkolsapaya J, Singhthong B, et al. Prediction, prevention, and mechanism of early (anaphylactic) antivenom reactions in victims of snake bites. Br Med J. 1986;292(6512):17-20. doi:10.1136/bmj.292.6512.17
194. Stone SF, Isbister GK, Shahmy S, Mohamed F, Abeysinghe C, Karunathilake H, et al. Immune response to snake envenoming and treatment with antivenom; complement activation, cytokine production and mast cell degranulation. PLoS Negl Trop Dis. 2013;7(7):e2326. doi:10.1371/journal.pntd.0002326
195. Laustsen AH, Engmark M, Milbo C, Johannesen J, Lomonte B, Gutiérrez JM, et al. From Fangs to Pharmacology: The Future of Snakebite Envenoming Therapy. Curr Pharm Des. 2016;22(34):5270-93. doi:10.2174/1381612822666160623073438
196. 196. León G, Segura A, Herrera M, Otero R, França FO, Barbaro KC, et al. Human heterophilic antibodies against equine immunoglobulins: assessment of their role in the early adverse reactions to antivenom administration. Trans R Soc Trop Med Hyg. 2008;102(11):1115-9. doi:10.1016/j.trstmh.2008.04.038
197. Silverstein AM. Clemens Freiherr von Pirquet: explaining immune complex disease in 1906. Nat Immunol. 2000;1(6):453-5. doi:10.1038/82691
198. Segura A, Herrera M, Villalta M, Vargas M, Gutiérrez JM, León G. Assessment of snake antivenom purity by comparing physicochemical and immunochemical methods. Biologicals. 2013;41(2):93-7. doi:10.1016/j.biologicals.2012.11.001
199. Otero-Patiño R, Cardoso JL, Higashi HG, Nunez V, Diaz A, Toro MF, et al. A randomized, blinded, comparative trial of one pepsin-digested and two whole IgG antivenoms for Bothrops snake bites in Uraba, Colombia. The Regional Group on Antivenom Therapy Research (REGATHER). Am J Trop Med Hyg. 1998;58(2):183-9. doi:10.4269/ajtmh.1998.58.183
200. Köhler G, Milstein C. Continuous cultures of fused cells secreting antibody of predefined specificity. J Immunol. 1975;174(5):2453-5.
201. Parray HA, Shukla S, Samal S, Shrivastava T, Ahmed S, Sharma C, et al. Hybridoma technology a versatile method for isolation of monoclonal antibodies, its applicability across species, limitations, advancement and future perspectives. Int Immunopharmacol. 2020;85:106639. doi:10.1016/j.intimp.2020.106639
202. Singh S, Kumar NK, Dwiwedi P, Charan J, Kaur R, Sidhu P, et al. Monoclonal Antibodies: A Review. Curr Clin Pharmacol. 2018;13(2):85-99. doi:10.2174/1574884712666170809124728
203. Courtenay-Luck NS, Epenetos AA, Moore R, Larche M, Pectasides D, Dhokia B, et al. Development of primary and secondary immune responses to mouse monoclonal antibodies used in the diagnosis and therapy of malignant neoplasms. Cancer Res. 1986;46(12 Pt 1):6489-93.
204. Basu K, Green EM, Cheng Y, Craik CS. Why recombinant antibodies - benefits and applications. Curr Opin Biotechnol. 2019;60:153-8. doi:10.1016/j.copbio.2019.01.012
205. Maverakis E, Kim K, Shimoda M, Gershwin ME, Patel F, Wilken R, et al. Glycans in the immune system and The Altered Glycan Theory of Autoimmunity: a critical review. J Autoimmun. 2015;57:1-13. doi:10.1016/j.jaut.2014.12.002
206. Townsend S, Finlay WJ, Hearty S, O'Kennedy R. Optimizing recombinant antibody function in SPR immunosensing. The influence of antibody structural format and chip surface chemistry on assay sensitivity. Biosens Bioelectron. 2006;22(2):268-74. doi:10.1016/j.bios.2006.01.010
207. Ma H, O'Kennedy R. The Structure of Natural and Recombinant Antibodies. Methods Mol Biol. 2015;1348:7-11. doi:10.1007/978-1-4939-2999-3_2
208. Bailon Calderon H, Yaniro Coronel VO, Cáceres Rey OA, Colque Alave EG, Leiva Duran WJ, Padilla Rojas C, et al. Development of Nanobodies Against Hemorrhagic and Myotoxic Components of Bothrops atrox Snake Venom. Front Immunol. 2020;11:655. doi:10.3389/fimmu.2020.00655
209. Luiz MB, Pereira SS, Prado NDR, Gonçalves NR, Kayano AM, Moreira-Dill LS, et al. Camelid Single-Domain Antibodies (VHHs) against Crotoxin: A Basis for Developing Modular Building Blocks for the Enhancement of Treatment or Diagnosis of Crotalic Envenoming. Toxins (Basel). 2018;10(4). doi:10.3390/toxins10040142
210. Souza JB, Cardoso R, Almeida-Souza HO, Carvalho CP, Correia LIV, Faria PCB, et al. Generation and In-planta expression of a recombinant single chain antibody with broad neutralization activity on Bothrops pauloensis snake venom. Int J Biol Macromol. 2020;149:1241-51. doi:10.1016/j.ijbiomac.2020.02.028
211. Campos LB, Pucca MB, Silva LC, Pessenda G, Filardi BA, Cerni FA, et al. Identification of cross-reactive human single-chain variable fragments against phospholipases A(2) from Lachesis muta and Bothrops spp venoms. Toxicon. 2020;184:116-21. doi:10.1016/j.toxicon.2020.05.027
212. Kaplon H, Reichert JM. Antibodies to watch in 2021. MAbs. 2021;13(1):1860476. doi:10.1080/19420862.2020.1860476
213. Eursakun S, Simsiriwong P, Ratanabanangkoon K. Studies on the fractionation of equine antivenom IgG by combinations of ammonium sulfate and caprylic acid. Toxicon. 2012;60(6):1022-9. doi:10.1016/j.toxicon.2012.07.005
214. Herrera M, Meneses F, Gutiérrez JM, León G. Development and validation of a reverse phase HPLC method for the determination of caprylic acid in formulations of therapeutic immunoglobulins and its application to antivenom production. Biologicals. 2009;37(4):230-4. doi:10.1016/j.biologicals.2009.02.020
215. Nudel BC, Perdoménico C, Iácono R, Cascone O. Optimization by factorial analysis of caprylic acid precipitation of non-immunoglobulins from hyperimmune equine plasma for antivenom preparation. Toxicon. 2012;59(1):68-73. doi:10.1016/j.toxicon.2011.10.014
216. Rojas G, Jiménez JM, Gutiérrez JM. Caprylic acid fractionation of hyperimmune horse plasma: description of a simple procedure for antivenom production. Toxicon. 1994;32(3):351-63. doi:10.1016/0041-0101(94)90087-6
217. Smith GP. Filamentous fusion phage: novel expression vectors that display cloned antigens on the virion surface. Science. 1985;228(4705):1315-7. doi:10.1126/science.4001944
218. Roncolato EC, Campos LB, Pessenda G, Costa e Silva L, Furtado GP, Barbosa JE. Phage display as a novel promising antivenom therapy: a review. Toxicon. 2015;93:79-84. doi:10.1016/j.toxicon.2014.11.001
219. Kumar R, Parray HA, Shrivastava T, Sinha S, Luthra K. Phage display antibody libraries: A robust approach for generation of recombinant human monoclonal antibodies. Int J Biol Macromol. 2019;135:907-18. doi:10.1016/j.ijbiomac.2019.06.006
220. Winter G, Griffiths AD, Hawkins RE, Hoogenboom HR. Making antibodies by phage display technology. Annu Rev Immunol. 1994;12:433-55. doi:10.1146/annurev.iy.12.040194.002245
221. Ledsgaard L, Kilstrup M, Karatt-Vellatt A, McCafferty J, Laustsen AH. Basics of Antibody Phage Display Technology. Toxins (Basel). 2018;10(6). doi:10.3390/toxins10060236
222. Frenzel A, Hust M, Schirrmann T. Expression of recombinant antibodies. Front Immunol. 2013;4:217. doi:10.3389/fimmu.2013.00217
223. Humphreys DP. Production of antibodies and antibody fragments in Escherichia coli and a comparison of their functions, uses and modification. Curr Opin Drug Discov Devel. 2003;6(2):188-96.
224. McKenna R, Lombana TN, Yamada M, Mukhyala K, Veeravalli K. Engineered sigma factors increase full-length antibody expression in Escherichia coli. Metab Eng. 2019;52:315-23. doi:10.1016/j.ymben.2018.12.009
225. Lakowitz A, Krull R, Biedendieck R. Recombinant production of the antibody fragment D1.3 scFv with different Bacillus strains. Microb Cell Fact. 2017;16(1):14. doi:10.1186/s12934-017-0625-9
226. Yang M, Zhu G, Korza G, Sun X, Setlow P, Li J. Engineering Bacillus subtilis as a Versatile and Stable Platform for Production of Nanobodies. Appl Environ Microbiol. 2020;86(8). doi:10.1128/aem.02938-19
227. Redwan E. The optimal gene sequence for optimal protein expression in Escherichia coli: principle requirements. Arab J Biotech. 2006;9(3):493-510.
228. Castro JM, Oliveira TS, Silveira CR, Caporrino MC, Rodriguez D, Moura-da-Silva AM, et al. A neutralizing recombinant single chain antibody, scFv, against BaP1, A P-I hemorrhagic metalloproteinase from Bothrops asper snake venom. Toxicon. 2014;87:81-91. doi:10.1016/j.toxicon.2014.05.017
229. Jia B, Jeon CO. High-throughput recombinant protein expression in Escherichia coli: current status and future perspectives. Open Biol. 2016;6(8). doi:10.1098/rsob.160196
230. Khow O, Suntrarachun S. Strategies for production of active eukaryotic proteins in bacterial expression system. Asian Pac J Trop Biomed. 2012;2(2):159-62. doi:10.1016/s2221-1691(11)60213-x
231. Porowińska D, Wujak M, Roszek K, Komoszyński M. [Prokaryotic expression systems]. Postepy Hig Med Dosw. 2013;67:119-29. doi:10.5604/17322693.1038351
232. Baumgarten T, Ytterberg AJ, Zubarev RA, de Gier JW. Optimizing Recombinant Protein Production in the Escherichia coli Periplasm Alleviates Stress. Appl Environ Microbiol. 2018;84(12). doi:10.1128/aem.00270-18
233. Hanyu Y, Kato M. High-yield expression of periplasmic single-chain variable fragments by solid Escherichia coli cultures. Biotechniques. 2022;72(1):29-32. doi:10.2144/btn-2021-0093
234. Turchetto J, Sequeira AF, Ramond L, Peysson F, Brás JL, Saez NJ, et al. High-throughput expression of animal venom toxins in Escherichia coli to generate a large library of oxidized disulphide-reticulated peptides for drug discovery. Microb Cell Fact. 2017;16(1):6. doi:10.1186/s12934-016-0617-1
235. Fan D, Liu C, Liu L, Zhu L, Peng F, Zhou Q. Large-scale gene expression profiling reveals physiological response to deletion of chaperone dnaKJ in Escherichia coli. Microbiol Res. 2016;186-187:27-36. doi:10.1016/j.micres.2016.03.001
236. Rivera-de-Torre E, Rimbault C, Jenkins TP, Sørensen CV, Damsbo A, Saez NJ, et al. Strategies for Heterologous Expression, Synthesis, and Purification of Animal Venom Toxins. Front Bioeng Biotechnol. 2021;9:811905. doi:10.3389/fbioe.2021.811905
237. Shanmugasundaram M, Pavlova NV, Pavlov AR, Lednev IK, Robb FT. Improved folding of recombinant protein via co-expression of exogenous chaperones. Methods Enzymol. 2021;659:145-70. doi:10.1016/bs.mie.2021.09.001
238. Bu D, Zhou Y, Tang J, Jing F, Zhang W. Expression and purification of a novel therapeutic single-chain variable fragment antibody against BNP from inclusion bodies of Escherichia coli. Protein Expr Purif. 2013;92(2):203-7. doi:10.1016/j.pep.2013.10.002
239. 239. Noguchi T, Nishida Y, Takizawa K, Cui Y, Tsutsumi K, Hamada T, et al. Accurate quantitation for in vitro refolding of single domain antibody fragments expressed as inclusion bodies by referring the concomitant expression of a soluble form in the periplasms of Escherichia coli. J Immunol Methods. 2017;442:1-11. doi:10.1016/j.jim.2016.11.014
240. Ahmad M, Hirz M, Pichler H, Schwab H. Protein expression in Pichia pastoris: recent achievements and perspectives for heterologous protein production. Appl Microbiol Biotechnol. 2014;98(12):5301-17. doi:10.1007/s00253-014-5732-5
241. Alvarenga LM, Zahid M, di Tommaso A, Juste MO, Aubrey N, Billiald P, et al. Engineering venom's toxin-neutralizing antibody fragments and its therapeutic potential. Toxins (Basel). 2014;6(8):2541-67. doi:10.3390/toxins6082541
242. Karbalaei M, Rezaee SA, Farsiani H. Pichia pastoris: A highly successful expression system for optimal synthesis of heterologous proteins. J Cell Physiol. 2020;235(9):5867-81. doi:10.1002/jcp.29583
243. Hou J, Tyo KE, Liu Z, Petranovic D, Nielsen J. Metabolic engineering of recombinant protein secretion by Saccharomyces cerevisiae. FEMS Yeast Res. 2012;12(5):491-510. doi:10.1111/j.1567-1364.2012.00810.x
244. Xie Y, Han X, Miao Y. An Effective Recombinant Protein Expression and Purification System in Saccharomyces cerevisiae. Curr Protoc Mol Biol. 2018;123(1):e62. doi:10.1002/cpmb.62
245. Kunert R, Reinhart D. Advances in recombinant antibody manufacturing. Appl Microbiol Biotechnol. 2016;100(8):3451-61. doi:10.1007/s00253-016-7388-9
246. Nieto-Taype MA, Garcia-Ortega X, Albiol J, Montesinos-Seguí JL, Valero F. Continuous Cultivation as a Tool Toward the Rational Bioprocess Development With Pichia Pastoris Cell Factory. Front Bioeng Biotechnol. 2020;8:632. doi:10.3389/fbioe.2020.00632
247. Vieira SM, da Rocha SLG, Neves-Ferreira A, Almeida RV, Perales J. Heterologous expression of the antimyotoxic protein DM64 in Pichia pastoris. PLoS Negl Trop Dis. 2017;11(7):e0005829. doi:10.1371/journal.pntd.0005829
248. Dumont J, Euwart D, Mei B, Estes S, Kshirsagar R. Human cell lines for biopharmaceutical manufacturing: history, status, and future perspectives. Crit Rev Biotechnol. 2016;36(6):1110-22. doi:10.3109/07388551.2015.1084266
249. Yin J, Li G, Ren X, Herrler G. Select what you need: a comparative evaluation of the advantages and limitations of frequently used expression systems for foreign genes. J Biotechnol. 2007;127(3):335-47. doi:10.1016/j.jbiotec.2006.07.012
250. Jenkins TP, Laustsen AH. Cost of Manufacturing for Recombinant Snakebite Antivenoms. Front Bioeng Biotechnol. 2020;8:703. doi:10.3389/fbioe.2020.00703
251. Chambers AC, Aksular M, Graves LP, Irons SL, Possee RD, King LA. Overview of the Baculovirus Expression System. Curr Protoc Protein Sci. 2018;91:5.4.1-5.4.6. doi:10.1002/cpps.47
252. Irons SL, Chambers AC, Lissina O, King LA, Possee RD. Protein Production Using the Baculovirus Expression System. Curr Protoc Protein Sci. 2018;91:5..1-5..22. doi:10.1002/cpps.45
253. Mishra V. A Comprehensive Guide to the Commercial Baculovirus Expression Vector Systems for Recombinant Protein Production. Protein Pept Lett. 2020;27(6):529-37. doi:10.2174/0929866526666191112152646
254. Possee RD, Chambers AC, Graves LP, Aksular M, King LA. Recent Developments in the Use of Baculovirus Expression Vectors. Curr Issues Mol Biol. 2020;34:215-30. doi:10.21775/cimb.034.215
255. Dubey KK, Luke GA, Knox C, Kumar P, Pletschke BI, Singh PK, et al. Vaccine and antibody production in plants: developments and computational tools. Brief Funct Genomics. 2018;17(5):295-307. doi:10.1093/bfgp/ely020
256. Fahad S, Khan FA, Pandupuspitasari NS, Ahmed MM, Liao YC, Waheed MT, et al. Recent developments in therapeutic protein expression technologies in plants. Biotechnol Lett. 2015;37(2):265-79. doi:10.1007/s10529-014-1699-7
257. Satheeshkumar PK. Expression of Single Chain Variable Fragment (scFv) Molecules in Plants: A Comprehensive Update. Mol Biotechnol. 2020;62(3):151-67. doi:10.1007/s12033-020-00241-3
258. Wang W, Yuan J, Jiang C. Applications of nanobodies in plant science and biotechnology. Plant Mol Biol. 2021;105(1-2):43-53. doi:10.1007/s11103-020-01082-z
259. Ko K. Expression of recombinant vaccines and antibodies in plants. Monoclon Antib Immunodiagn Immunother. 2014;33(3):192-8. doi:10.1089/mab.2014.0049
260. Liénard D, Sourrouille C, Gomord V, Faye L. Pharming and transgenic plants. Biotechnol Annu Rev. 2007;13:115-47. doi:10.1016/s1387-2656(07)13006-4
261. Poisot T, Péquin B, Gravel D. High-throughput sequencing: a roadmap toward community ecology. Ecol Evol. 2013;3(4):1125-39. doi:10.1002/ece3.508
262. Arivaradarajan P, Misra G. Omics Approaches, Technologies And Applications. 1 ed. Singapore: Springer; 2018. 1-5 p.
263. Calvete JJ. Snake venomics: from the inventory of toxins to biology. Toxicon. 2013;75:44-62. doi:10.1016/j.toxicon.2013.03.020
264. León G, Segura A, Gomez A, Hernández A, Navarro D, Villalta M, et al. Industrial Production and Quality Control of Snake Antivenoms. In: Gopalakrishnakone P, Calvete J, editors. Venom Genomics and Proteomics. Dordrecht: Springer; 2016. p. 425-50.
265. León G, Vargas M, Segura Á, Herrera M, Villalta M, Sánchez A, et al. Current technology for the industrial manufacture of snake antivenoms. Toxicon. 2018;151:63-73. doi:10.1016/j.toxicon.2018.06.084
266. Patra A, Banerjee D, Dasgupta S, Mukherjee AK. The in vitro laboratory tests and mass spectrometry-assisted quality assessment of commercial polyvalent antivenom raised against the 'Big Four' venomous snakes of India. Toxicon. 2021;192:15-31. doi:10.1016/j.toxicon.2020.12.015
267. Yadav D, Tanveer A, Malviya N, Yadav S. Chapter 1 - Overview and Principles of Bioengineering: The Drivers of Omics Technologies. In: Barh D, Azevedo V, editors. Omics Technologies and Bio-Engineering: Academic Press; 2018. p. 3-23.
268. Hubank M. Gene expression profiling and its application in studies of haematological malignancy. Br J Haematol. 2004;124(5):577-94. doi:10.1111/j.1365-2141.2004.04821.x
269. Rodrigues RS, Boldrini-França J, Fonseca FP, de la Torre P, Henrique-Silva F, Sanz L, et al. Combined snake venomics and venom gland transcriptomic analysis of Bothropoides pauloensis. J Proteomics. 2012;75(9):2707-20. doi:10.1016/j.jprot.2012.03.028
270. Freitas-de-Sousa LA, Amazonas DR, Sousa LF, Sant'Anna SS, Nishiyama MY, Jr., Serrano SM, et al. Comparison of venoms from wild and long-term captive Bothrops atrox snakes and characterization of Batroxrhagin, the predominant class PIII metalloproteinase from the venom of this species. Biochimie. 2015;118:60-70. doi:10.1016/j.biochi.2015.08.006
271. Amorim FG, Morandi-Filho R, Fujimura PT, Ueira-Vieira C, Sampaio SV. New findings from the first transcriptome of the Bothrops moojeni snake venom gland. Toxicon. 2017;140:105-17. doi:10.1016/j.toxicon.2017.10.025
272. Laustsen AH, Engmark M, Clouser C, Timberlake S, Vigneault F, Gutiérrez JM, et al. Exploration of immunoglobulin transcriptomes from mice immunized with three-finger toxins and phospholipases A(2) from the Central American coral snake, Micrurus nigrocinctus. PeerJ. 2017;5:e2924. doi:10.7717/peerj.2924
273. Schmidt A, Forne I, Imhof A. Bioinformatic analysis of proteomics data. BMC Syst Biol. 2014;8 Suppl 2(Suppl 2):S3. doi:10.1186/1752-0509-8-s2-s3
274. Gupta N, Tanner S, Jaitly N, Adkins JN, Lipton M, Edwards R, et al. Whole proteome analysis of post-translational modifications: applications of mass-spectrometry for proteogenomic annotation. Genome Res. 2007;17(9):1362-77. doi:10.1101/gr.6427907
275. Twyman R. Principles of Proteomics. 2 ed. Abingdon, UK: Garland Science; 2013. 1-148 p.
276. Banks RE, Dunn MJ, Hochstrasser DF, Sanchez JC, Blackstock W, Pappin DJ, et al. Proteomics: new perspectives, new biomedical opportunities. Lancet. 2000;356(9243):1749-56. doi:10.1016/s0140-6736(00)03214-1
277. Tashima AK, Zelanis A. Snake Venom Peptidomics. In: Gopalakrishnakone P, Calvete JJ, editors. Venom Genomics and Proteomics. Dordrecht: Springer Netherlands; 2016. p. 317-31.
278. Becker CH, Bern M. Recent developments in quantitative proteomics. Mutat Res. 2011;722(2):171-82. doi:10.1016/j.mrgentox.2010.06.016
279. Sanz L, Quesada-Bernat S, Ramos T, Casais ESLL, Corrêa-Netto C, Silva-Haad JJ, et al. New insights into the phylogeographic distribution of the 3FTx/PLA(2) venom dichotomy across genus Micrurus in South America. J Proteomics. 2019;200:90-101. doi:10.1016/j.jprot.2019.03.014
280. Galizio NDC, Serino-Silva C, Stuginski DR, Abreu PAE, Sant'Anna SS, Grego KF, et al. Compositional and functional investigation of individual and pooled venoms from long-term captive and recently wild-caught Bothrops jararaca snakes. J Proteomics. 2018;186:56-70. doi:10.1016/j.jprot.2018.07.007
281. Sousa LF, Portes-Junior JA, Nicolau CA, Bernardoni JL, Nishiyama MY, Jr., Amazonas DR, et al. Functional proteomic analyses of Bothrops atrox venom reveals phenotypes associated with habitat variation in the Amazon. J Proteomics. 2017;159:32-46. doi:10.1016/j.jprot.2017.03.003
282. De Sousa CS, Hassan SS, Pinto AC, Silva WM, De Almeida SS, De Castro Soares S, et al. Chapter 1 - Microbial Omics: Applications in Biotechnology. In: Barh D, Azevedo V, editors. Omics Technologies and Bio-Engineering. 2. 1 ed: Academic Press; 2018. p. 3-20.
283. Valente RH, Nicolau CA, Perales J, Neves-Ferreira AGdC. Snake Venom Proteopeptidomics: What Lies Behind the Curtain. In: Gopalakrishnakone P, Calvete JJ, editors. Venom Genomics and Proteomics. Dordrecht: Springer Netherlands; 2016. p. 333-65.
284. Jorge RJ, Monteiro HS, Gonçalves-Machado L, Guarnieri MC, Ximenes RM, Borges-Nojosa DM, et al. Venomics and antivenomics of Bothrops erythromelas from five geographic populations within the Caatinga ecoregion of northeastern Brazil. J Proteomics. 2015;114:93-114. doi:10.1016/j.jprot.2014.11.011
285. Quintana-Castillo JC, Vargas LJ, Segura C, Estrada-Gómez S, Bueno-Sánchez JC, Alarcón JC. Characterization of the Venom of C. d. cumanesis of Colombia: Proteomic Analysis and Antivenomic Study. Toxins (Basel). 2018;10(2). doi:10.3390/toxins10020085
286. Ogonah OW, Polizzi KM, Bracewell DG. Cell free protein synthesis: a viable option for stratified medicines manufacturing? Current Opinion in Chemical Engineering. 2017;18:77-83. doi:https://doi.org/10.1016/j.coche.2017.10.003
287. Zemella A, Thoring L, Hoffmeister C, Kubick S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. Chembiochem. 2015;16(17):2420-31. doi:10.1002/cbic.201500340
288. Joyce AR, Palsson B. The model organism as a system: integrating 'omics' data sets. Nat Rev Mol Cell Biol. 2006;7(3):198-210. doi:10.1038/nrm1857
289. Mochida K, Shinozaki K. Advances in omics and bioinformatics tools for systems analyses of plant functions. Plant Cell Physiol. 2011;52(12):2017-38. doi:10.1093/pcp/pcr153
290. Nalluri JJ, Barh D, Azevedo V, Ghosh P. Chapter 13 - Bioinformatics and Systems Biology in Bioengineering. In: Barh D, Azevedo V, editors. Omics Technologies and Bio-Engineering: Academic Press; 2018. p. 223-43.
291. De Arco-Rodríguez B, Montealegre-Sánchez L, Solano-Redondo L, Castro-Herrera F, Ortega JG, Castillo A, et al. Phylogeny and toxicological assessments of two Porthidium lansbergii lansbergii morphotypes from the caribbean region of Colombia. Toxicon. 2019;166:56-65. doi:10.1016/j.toxicon.2019.05.010
292. Neri-Castro E, Lomonte B, Valdés M, Ponce-López R, Bénard-Valle M, Borja M, et al. Venom characterization of the three species of Ophryacus and proteomic profiling of O. sphenophrys unveils Sphenotoxin, a novel Crotoxin-like heterodimeric β-neurotoxin. J Proteomics. 2019;192:196-207. doi:10.1016/j.jprot.2018.09.002
293. Simizo A, Kitano ES, Sant'Anna SS, Grego KF, Tanaka-Azevedo AM, Tashima AK. Comparative gender peptidomics of Bothrops atrox venoms: are there differences between them? J Venom Anim Toxins Incl Trop Dis. 2020;26:e20200055. doi:10.1590/1678-9199-jvatitd-2020-0055
294. Engmark M, Lomonte B, Gutiérrez JM, Laustsen AH, De Masi F, Andersen MR, et al. Cross-recognition of a pit viper (Crotalinae) polyspecific antivenom explored through high-density peptide microarray epitope mapping. PLoS Negl Trop Dis. 2017;11(7):e0005768. doi:10.1371/journal.pntd.0005768
295. Madrigal M, Alape-Girón A, Barboza-Arguedas E, Aguilar-Ulloa W, Flores-Díaz M. Identification of B cell recognized linear epitopes in a snake venom serine proteinase from the central American bushmaster Lachesis stenophrys. Toxicon. 2017;140:72-82. doi:10.1016/j.toxicon.2017.10.022
296. Molina Molina DA, Guerra-Duarte C, Naves de Souza DL, Costal-Oliveira F, Ávila GR, Soccol VT, et al. Identification of a linear B-cell epitope in the catalytic domain of bothropasin, a metalloproteinase from Bothrops jararaca snake venom. Mol Immunol. 2018;104:20-6. doi:10.1016/j.molimm.2018.10.019
297. Ramos HR, Junqueira-de-Azevedo Ide L, Novo JB, Castro K, Duarte CG, Machado-de-Ávila RA, et al. A Heterologous Multiepitope DNA Prime/Recombinant Protein Boost Immunisation Strategy for the Development of an Antiserum against Micrurus corallinus (Coral Snake) Venom. PLoS Negl Trop Dis. 2016;10(3):e0004484. doi:10.1371/journal.pntd.0004484
298. Schneider FS, de Almeida Lima S, Reis de Ávila G, Castro KL, Guerra-Duarte C, Sanchez EF, et al. Identification of protective B-cell epitopes of Atroxlysin-I: A metalloproteinase from Bothrops atrox snake venom. Vaccine. 2016;34(14):1680-7. doi:10.1016/j.vaccine.2016.02.035
299. de Castro KLP, Lopes-de-Souza L, de Oliveira D, Machado-de-Ávila RA, Paiva ALB, de Freitas CF, et al. A Combined Strategy to Improve the Development of a Coral Antivenom Against Micrurus spp. Front Immunol. 2019;10:2422. doi:10.3389/fimmu.2019.02422
300. P DVdM, de Almeida Lima S, Araújo P, Medina Santos R, Gonzalez E, Alves Belo A, et al. Immunoprotection against lethal effects of Crotalus durissus snake venom elicited by synthetic epitopes trapped in liposomes. Int J Biol Macromol. 2020;161:299-307. doi:10.1016/j.ijbiomac.2020.05.171
301. Lopes-de-Souza L, Costal-Oliveira F, Stransky S, Fonseca de Freitas C, Guerra-Duarte C, Braga VMM, et al. Development of a cell-based in vitro assay as a possible alternative for determining bothropic antivenom potency. Toxicon. 2019;170:68-76. doi:10.1016/j.toxicon.2019.09.010
Received: May 25, 2022 / Accepted: October 22, 2022 / Published:15 November 2022
Citation: Romo, E.; Torres, M.; Martin-Solano, S. Current situation of snakebites envenomation in the Neotropics: Biotechnology, a versatile tool in the production of antivenoms. Revis Bionatura 2022;7(4) 54. http://dx.doi.org/10.21931/RB/2022.07.04.54