2023.08.01.87
Files > Volume 8 > Vol 8 No 1 2023
Estimating genetic parameters of maize hybrids and parents under different plant densities (Combining ability for yield and some other traits for maize Zea mays L.)

Banan H. Hadi1* , Wajeeha A.Hassan2 , Zainab K. Alshugeairy3 , Faez F. Alogaidi4
1 Crop Sciences Department, College of Agriculture Engineering Sciences, University of Baghdad, Iraq.
2 Crop Sciences Department, College of Agriculture Engineering Sciences, University of Baghdad, Iraq. [email protected].
3 Crop Sciences Department, College of Agriculture Engineering Sciences, University of Baghdad, Iraq. [email protected]
4 Crop Sciences Department, College of Agriculture Engineering Sciences, University of Baghdad, Iraq. [email protected].
*Correspondence: : [email protected].
Available from: http://dx.doi.org/10.21931/RB/2023.08.01.87
ABSTRACT
A field experiment was carried out in the fields of the Field Crops Department - Faculty of Agricultural Engineering Sciences. The study included five inbred lines (ZM43W (ZE), ZM60, ZM49W3E, ZM19, CDCN5), given numbers 1, 2, 3, 4 and 5) to study the hybrid vigor and both general and special combing ability (GCA, SCA) of the half diallel mating method, for the spring and fall seasons (2016). The genetic analysis shows that all crosses gave a positive hybrid vigor for grain yield per unit area at the two population densities. the highest value is 116.20% for cross (3´5 )at low density, and 89.22% for cross( 1´4 )at high density. The hybrid vigor for all crosses is positive at two densities for dry matter yield, crop growth rate and ears weight. The highest value is 81.31%, 96.30% and 131.45% at high density for these traits for the cross (1´2), respectively. Also, this cross gave the highest value at high density for grain yield per plant (170.61%) and (85.43%) for no. of grain plant-1. The general combining ability in two densities for all studied traits. The highest positive value was (48.949) for parent 3 at low density. All values of s2sca are more than values of s2gca, and all values of s2D are more than all values of s2A. For this, all h2n.s. were low. It ranges from 1.88% for the crop growth rate to 18.82% for no. of rows ear-1 at low density and between -0.38 for the crop growth rate to 41.42 for 300-grain weight at high density. Because the values of s2D are higher than values of s2A, the values of the ratio of s2gca/s2gsca were less than one, while the value of were mor than one. This indicates that all these traits are influenced by dominance genes, and the importance of the non-additive gene action and its large contribution to the inheritance of these traits.
Keywords: maize, combining ability, heritability, genetic parameters.
INTRODUCTION
The primary aim of plant breeding is identifying or finding a superior germplasm pool, selecting a superior individual, and developing a superior variety from the selection. Reported1 that knowing how genes act and interact will determine which breeding system optimizes gene action more efficiently and will elucidate the role of the breeding systems in the diallel mating systems is good design. Estimates of GCA and SCA effects are appropriate and instrumental genetic parameters of the parents and their crosses2. Estimating genetic effects is appropriate for more diallel mating systems, but investigators often desire to extend estimation to include genetic components of variance and heritability. It is necessary to obtain information on the nature of the combining ability of the parents, to know their behavior and performance in the crosses, to select the best ones for crossing, and to diagnose hybrids or promising varieties3. The analysis of both general and specific combining ability is an important tool for selecting the desired parent and obtaining information about the rapture and magnitude of the effect of the gene-controlled quantitative traits4. Heterosis occurs when the crosses exceed the parents' average because of non-additive genetic effects. There were different opinions about the relative importance of additive and non-additive variance in maize populations. Interactions of alleles either within the same loci or among loci were necessary. Found5 that estimates of additive genetic variance were two to four times greater than estimates due to dominance deviations in maize; hence, selection should be effective. showed6 theoretically that selection among single crosses would be twice as effective as selection among double crosses if only additive genetic effect were considered. If non-additive effects (dominance and epistasis) were important in the genetic variances among types of hybrids, the advantage of selection among single crosses rather than among double crosses would be even greater. Selection, natural or humane, was based on continuous variation with the replacement of unfavorable alleles by more favorable alleles and the accumulation of modifiers in support of the more important alleles2. The role of deleterious genes in the expression of heterosis in maize is decisive. Deleterious genes are preserved in large numbers in maize inbred lines through selection for combining ability, which is why hybrids are much more productive than inbred lines. The more harmful genes are replaced by favorable additive alleles, the more the yielding capacity of inbred lines improves, and heterozygosity7 favors the less the yield in F1. Production of vigorous inbred lines that approach or out-yield hybrid productivity is feasible only when selection for combining ability is replaced by selection for inbred line performance per se so that deleterious genes can be effectively removed. illustrate8 that the inbred L3 had positive gca. The cross (L3´T8 ) had the highest positive hybrid vigor for grain yield (98.61%) and sca (18.98). The value of additive variation for inbred was more than for tester for all traits. The dominant gene action for inbred was lower than additive, but it was close to additive variation for testers. s2gca and genetic variation for inbred were superior to testers for most traits. The average degree of dominance was more than one, except grain weight was less than one. Found9 that the inbred 2 had positive GCA. All parents and crosses had a positive variance for yield. The mean square for SCA was more than GCA, and therefore the s2GCA was less, which made s2A less than s2D, so s2GCA/s2SCA became less than one, and the degree of dominance was more than one, making h2.n.s. very little. Thus, the trait was governed by non-additive gene action10. found11 that the genetic analysis shows important both additive and dominant effects to heredity of the yield and yield components throw significant the GCA, SCA. The dominance effect was more important than the additive effect. The GCA /SCA is less than one and s2D higher than s2A. The degree of dominance is more than one, and high broad sense heritability and low, narrow sense heritability for all traits. The results12 obtained indicated that all estimates of additive (V.A.) and dominance (V.D.) variance were significant for all traits except V.A. for no. of kernel row-1. The magnitude of V.A. was consistently larger than that of V.D. for all traits except for grain yield, where V.D. values were larger than V.A. values. High narrow sense heritability estimates were detected for no. of kernel row-1. Moderate h2n.s. estimates were obtained for 100 kernel weights, while the estimate was for grain yield.
MATERIALS AND METHODS
A field experiment was carried out in the fields of the Field Crops Department - Faculty of Agricultural Engineering Sciences. The study included five inbred lines (ZM43W (ZE), ZM60, ZM49W3E, ZM19, CDCN5), given numbers 1, 2, 3, 4 and 5) to study the hybrid vigor and both general and unique combing ability (GCA, SCA) of the half diallel mating method, for the spring and fall seasons of 2016. The soil was prepared as recommended. Triple superphosphate fertilizer 46% P2O5 was added by 200 kg and urea fertilizer 46% by 300 kg N ha-1 in three batches when planting and two months after adding the first batch and upon flowering. In the first season, the seeds of the inbred lines were planted on 4/4/2016 in furrows; the length of the furrows was 7 meters, the distance between one furrow and the other was 0.75 meters, and the distance between one plant and another was 25cm. When the plants reached the flowering stage, the female inflorescences were wrapped a day before pollination before the release of the silk, and the male inflorescences were wrapped a day before pollination. Half diallel crosses were performed between inbred lines. After the maturity of the plants, they were harvested separately to be planted in the fall season. In the second season, the seeds of the resulting crosses were planted 10 crosses in addition to the five parents to evaluate their performance under two plant densities of 50 and 70 thousand plants hectare-1 by designing randomized complete block ( RCBD ) and in the arrangement of the spilled plot, as the plant densities represented the main plots and the genotypes described the secondary plots. On furrows, the length of the furrow is 6 m. The distance between one furrow and another is 0.75, and between one plant and another is 0.266 and 0.1904 m for the two densities mentioned in sequence. At the end of the season, random samples were taken from 5 plants to measure the following traits: number of rows per ear, number of grains per row, number of grains per ear, the weight of 300 grains, ears weight, grain yield per plant, grain yield for area unit, total dry weight and crop growth rate, Statistical analysis was done using the Genstate program hybrid vigor the half diallel analysis of the traits that showed significant differences were calculated by statistical analysis and according to the fourth method, the Fixed Model of Griffng's analysis to estimate the general combining ability (GCA) and the specific combining ability ( SCA ) according to what14 which divides the mean squares of the genotypes into the mean squares of general combining ability (GCA) and mean squares of specific combining ability (SCA ) According to the following mathematical model:


RESULTS
Hybrid vigor
Hybrid vigor or heterosis, the increased performance of the hybrid progenies compared to their homozygous parents, reaches high levels in maize with certain combinations of complementary heterotic groups.
Number of rows for the ear
Table 1 shows that the hybrid vigor for crosses significantly differed at the two densities. The cross (1´2), (1´4), (3´4) and (3´5) showed negative hybrid vigor at the low density (50000 p/h.) and positive at the high density (70000 p/h.). Cross (1´3) and (2´4) have positive hybrid vigor at the two densities, while cross( 1´5),( 2´3), and (4´5) give negative hybrid vigor at two densities. The cross (2´5) had a positive hybrid vigor at low densities and negative at high.
.Number of grains per row
All crosses had positive values for hybrid vigor at two densities, except cross 2´3 was negative at high density.
Number of grains per plant
All values of hybrid vigor for all crosses were positive at two densities. The highest value was 62,98% for the cross( 4´5) at low density and 85.43% for the cross (1´2) at high density, Table 1. The positive value of hybrid vigor for crosses means a tendency to increase the no. of grains of the plant. We note that these two crosses possessed a SCA positive and high. This is due to the difference between their parents in the GCA, as one was positive and the other was negative.
The weight of ear
The hybrid vigor values differed significantly between crosses at the two densities. All values were positive in the direction of increasing the weight of the ear of maize. Cross (2´4) and ( 4´5) achieved the highest value (58.24% and 55.34%) at the low density but decreased at high density (14.90% and 14.47%). That is, the gene expression for these crosses appears at low density (did not tolerate the high density). Cross 2 achieved the highest value' (131.45%) at high density and low value' at the low-density table 1.
The weight of 300 grains
Table 1 shows that 7 crosses gave a negative hybrid vigor at low density, the highest value -21.05 for the cross (1´5). The cross (2´3), (2´4) and (3´4) gave positive value for hybrid vigor. in high density, 6 crosses (1´3), ( 2´3), ( 2´4), (3´4), ( 3´5) and( 4´5) gave a negative value and cross (1´2), (1´4), (1´5), and 2´5) gave positive value.
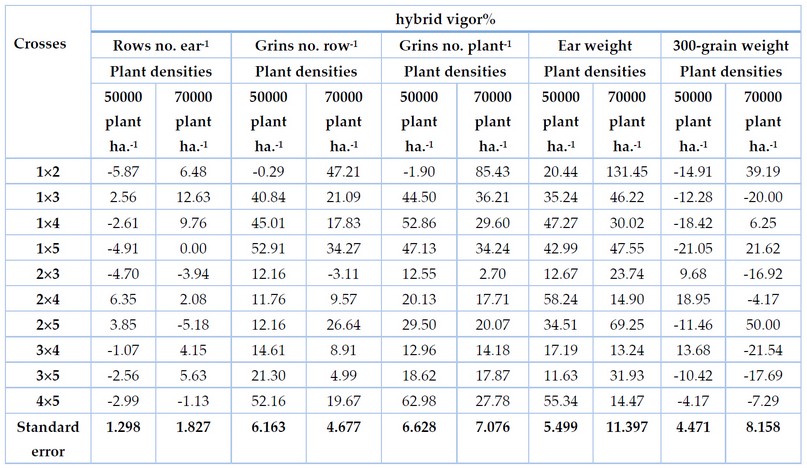
Table 1. Percentage of hybrid vigor of maize crosses for yield component traits in different plant densities for fall season 2016.
Grain yield per plant
All crosses achieved positive values for hybrid vigor at low density. Values ranged from 18.02% for the cross (1´2) to 80.09% for the cross (2´4). The positive value of the hybrid vigor indicates the cross's ability to increase the plant's yield. In high density, two of the ten crosses gave a negative value of hybrid vigor. The highest positive value is 170.61% for the cross( 1´2), which gave the lowest positive value at the low-density Table 2.
Grain yield per hectar
Table 2 shows that all crosses gave positive values for hybrid vigor at two densities in the direction to increase the yield.
Dry matter yield
Also, all values of hybrid vigor for all crosses were positive in increasing the yield of dry matter of the plant of maize.
The growth rate of plant
Table 2 shows that all values of the growth rate of the plant were positive at two densities. It ranged between 65.57% for cross (1´4) to 29.73 for cross (3´5) at low density. In high density, it ranged between 96.30% for cross (1´2) to 12.99% for cross (2´3).
When the value of the hybrid vigor is positive and high. This means the occurrence of the trait under the influence of the overdominance of the genes of the highest parents.

Table 2. Percentage of hybrid vigor of maize crosses for yield and growth criteria traits in different plant densities for fall season 2016.
The GCA effect
The number of rows per ear
Table 3 illustrates that parents 1 and 3 had a positive GCA at low density and a negative GCA at high density. Parents 2 and 4 had a negative GCA at low density and a positive GCA at high density. Parent 5 had a positive GCA at the two densities.
The number of grains per row
Parents differed significantly in the GCA. The parent 1 and 5 had a negative GCA at low density and positive at high density, while the parents 2 and 3 were the opposite, they were positive at low density and negative at high density. As for parent 4, it was positive at the tow densities.
Number of grain per plant
Three of parents have negative GCA at the both density. While parent 3 was positive, as for parent 5 was negative at low density and positive at high table 3.
The weight of ear
The table 3 illustrate that the parent 1,2and 4 had negative GCA at the two densities. The parents 3 had positive GCA at tow densities While the parent 5 had positive value GCA for high density and negative for low density.
Weight of 300 grains
The table 3 illustrate that the parent 2 and 5 had negative GCA at the two densities. the parents 1 and 4 have positive GCA at low density and negative at high density .the parent3 have positive GCA at the tow densities.

Table 3. The effects of general combining ability of maize parents for yield component traits in different plant densities for fall season 2016.
The yield of plant
Parent 1 and 2 showed a negative GCA in both densities. Parent 3 showed positive GCA at tow densities, while parent 4 was positive at low density and negative GCA at high density. But the parent 5, was negative at low density and positive at high table 4.
Yield per unit area
Parents differed significantly in the GCA at two densities. parents 1 and 4 showed negative GCA at two densities. parents 3 and 5 showed positive GCA at two densities. While the parent 2 showed negative at low density and positive at high table 4.
Dry matter
Two parents 1 and 2 showed negative GCA at both densities, parent 3 was positive at two densities, while parent 4 was positive at low densities and positive in high. But the parent 5 was negative at low and positive at high densities table 4.
The growth rate of plant
The parent 1 have negative GCA at low density and positive at high (Table 4). Parent 2 have negative GCA at two densities, opposite of parent 3 was positive at two densities. Parents 4 and 5 were positive at low density and negative at high density.

Table 4. The effects of general combining ability of maize parents for yield and growth criteria traits in different plant densities for fall season 2016.
Specific combining ability
The sca measures the effects of non-additive genes action. When its value is high, this means a great agreement between parents of the yield.
Number of rows
Four crosses (1´3), (1´4), (2´4), and (3´4) have positive SCA at two densities, the cross (1´5) and 2´3 have negative SCA at two densities. The cross 1´2, 3´5, have negative SCA at low densities and positive SCA at high. The cross 2´5 and 4´5 have positive SCA at low density and negative at high (Table 5).
Number of grains per row
Table 5 showed that seven crosses of maize have positive SCA at two density, while one cross only have negative SCA at both density, and one is negative at the low density and positive at high density, and one is opposite it is positive at low density and negative at high density.
Number of grains per plant
There are eight crosses of maize showed positive SCA at the both densities. While the cross (2´3) was negative at the two densities. The cross (1´2) was negative at the low density and positive at high density.
The weight of the ear
Table 5 showed that all crosses showed positive SCA at the two densities, except the cross (1´2) was negative at the low density and positive at the high density.
Weight of 300 grains
Table 3 showed that five crosses have negative SCA at the low density, and positive SCA the high density. while three crosses have positive SCA at low density and negative SCA at high density. Only two crosses (1´3) and (4´5) have negative SCA at both densities.

Table 5. The effects of specific combining ability of maize crosses for yield and components traits in different plant densities for fall season 2016.
Yield of plant
Eight crosses have positive SCA at both densities (Table 6). The highest value is 70.67 for cross (1´3) and 58.76 for cross 2´4 at the low density. The highest value is 76.32 for cross (2´5) followed by 60.70 for cross (1´2). When the value of SCA is high, it means a great agreement between the parents of cross for the yield. Two of crosses (2´3) and (3´4) have appositive SCA at low density and negative in high density.
The yield of unit area
All crosses showed positive SCA at the two densities. The highest value is 3.37 and 2.40 for cross (3´5) at both densities respectively, followed by 2.02 and 2.79 for cross (1´2) at both density respectively table 6.
.Dry matter for maize plant
Table 6 indicate that all crosses have positive SCA at both densities, except the cross (3´4 ) is negative in the high density. The highest value is 81.00 for cross (1´3) at low density. followed by 50.62 for cross (2´4). In the high density, the highest value is 73.87 for cross (1´2) followed by 52.30 and 50.97 for cross (1´3) and (2´5 ), respectively.
The growth rate of maize plant
As in the previous trait, all crosses showed a positive SCA at the two densities, except the cross (3´4) is negative in high density Table 6.

Table 6. The effects of specific combining ability of maize crosses for yield and growth criteria traits in different plant densities for fall season 2016.
Genetic parameters
Low density
Table 7 showed that all values of non-additive variance (s2D) were high than the values of Additive variance (s2A) for all studies traits. Hence the values of specific combining ability variance (s2sca) were higher than the values of general combining ability variance (s2gca). The highest value was 16496.87 for the no. of grains per plant, followed by 3469. 95 for der matter of maize plant and 2572.43 for grain yield of plant. While the highest s2gca/s2sca was 05488 for trait no. of rows per ear, but the lowest value was 0.0101 for the growth rate followed by 0.0176 for dry matter. All values of broad sense heritability are high, due to the approach of the genetic variance values to the phenotypic variance values. Except the value of broad sense heritability for no. of rows per ear is moderate. The values of narrow sense heritability ranged from 1.88% for the trait growth rate of the crop to 18.82% for the no. of rows per ear, followed by the weight of 300 grains, for which the heritability value reached 15.98%. The values of the average degree of dominance for all the studied traits were more than one, indicating that these traits are under the effect of over dominance of gene's. It also shows the importance of the non-additive gene action and its large contribution to the inheritance of these traits.
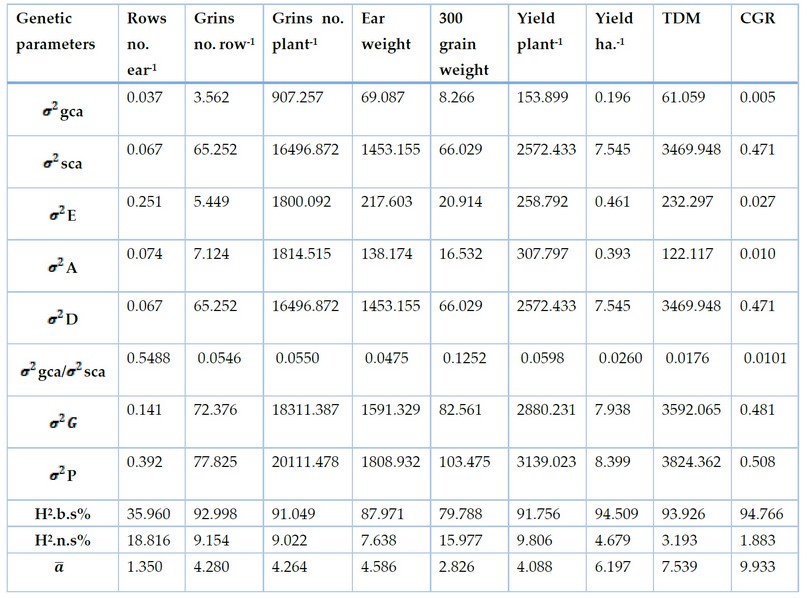
Table 7. Genetic parameters of maize parents and hybrids under low plant density ( 50000 )plants hectare-1 for yield and components traits for fall season 2016.
Genetic parameters
High density
As in the previous table, we find that all the values of the variance of the specific combining ability (s2sca) are higher than the values of the conflict of the general combining power (s2gca) of all the studied traits Table 8. As well, all matters of the variance of non-additive gene action (s2D) are higher than the variance of additive (s2A). The higher value of s2D is 16496.87 for trait no. of grains per plant, followed by 3469.95 for trait dry matter of maize plant, and 2572.43 for trait grains yield of the plant. While the higher value of s2A is 1814.52 for trait no. of grains per plant too. For this, all s2gca/s2sca values were less than one, indicating the importance of non-additive gene action in the inheritance of these traits and the additive gene effect. Table 8, shows that all the values of genetic variance (s2g) are close to the importance of phenotypic variance (s2p), so all values of the broad sense heritability were high. It ranged from 82.07% for a trait of the no. of rows per ear to 94.83% for a trait 300 grain. The values of the narrow sense heritability ranged between ( 0.38%) for the crop growth rate (due to the low value of the variance of additive s2A) to 41.42% for the weight of 300 grains. All matters of the average degree of dominance for all studied traits were more than one. This indicates that all these traits are under the influence of dominance genes and indicates the importance of the non-additive gene action and its large contribution to the inheritance of these traits.

Table 8. Genetic parameters of maize parents and hybrids under high plant density ( 70000 )plants hectare-1 for yield and components traits for fall season 2016.
DISCUSSION
Three main genetic mechanisms have been proposed to explain heterosis: dominance, overdominance and positive epistatic interaction14. Moreover, the highest percentage of heterosis for grain per ear over better parent was observed by15 the cross P2´P3. However, positive general combining ability indicates that these parents contribute to improving the trait and transmit the additive gene effect for high grain yield to its crosses through its contribution to the inheritance of the trait towards increasing the yield. So, these parents can be used in hybridization programs to improve the yield and increase yield efficiency by selecting a superior plant in grain yield. The magnitude of GCA was higher than the sca in all the cases indicating that additive gene action was more important than non-additive in the inheritance of this traits16. The values of GCA and SCAC for grain yield and its component are highly significant. The dominance gene effect for grain yield and no. of grains per row is more significant, while the additive gene action is more important for grain weight17. The additive gene effect is more important than the non-additive in inheriting ear weight18. GCA and SCA mean squares were highly significant for all traits, but for no. of rows ear-1 SCA mean square was significant19. Significant GCA and SCA variance were observed for all traits except ear height. The non-additive variation of the studied traits of the maize plant was high, making the ratio s2gca/s2sca less than one), indicating the importance of non-additive genetic influences and the additive effect of genes in the inheritance of the grain yield. This result agrees with what both of found10,20 that the non-additive gene action controlled the outcome. The additive genetic variance was preponderant for grains per ear and 1000-grain weight24. Several investigators reported that additive gene action was responsible for the inheritance of grain yield and most of its contributing traits21. However, it reported22,23 that the non-additive gene action was more important in maize inheritance grain yield and most other agronomic traits. While reported20,24 that both additive and non-additive gene effects were important in the genetic expression of maize yield and its contributing features. The s2gca/s2sca for all traits were less than one, except no. of rows ear-1 was more than one25.
CONCLUSION
We can conclude that the parents differ in their performance and general and specific combining ability. Some of the parents had a positive general combining ability at the two densities, while others showed different general combining abilities of each density. Some parents showed a high special combining ability between them, which medicates the possibility of using them in breeding programs to produce superior hybrids or synthetic varieties.
REFERENCES
1. Janick, J. Plant Breeding Reviews. Purdue Univ. John Wiley & Sons, Inc. vol.1997.; 15 P. 316.
2. Hallauer, A.R. History, contribution and future of quantitative genetics in plant breeding. Lessons from maize. Crop Sci. 2007.;47(3): 4-19.
3. Bello, O.B.; Olaoye, G. Combining ability for maize grain yield and other agronomic characters in typical southern guinea savanna ecology of Nigeria. Afr. J. Biotechnology. 2009.; 8 2518-2522.
4. Basbag, S.; Ekinci, R.; Gencer, O. Combining ability and heterosis for earliness characters in line´tester population of Gossypium hirsarum L. Hereditas, 2007.; 144: 185-190.
5. Hallauer, A.R.; Miranda, J.B. Quantitative Genetic in Maize Breeding. 2nd ed. Iowa State Univ. Press, Ames, IA. 1988.;
6. Cockerham, C.C. Implications of genetic variance in a hybrid breeding program. Crop Sci. 1961.; 1: 47-52.
7. Fasoulas, A.C.; Principles of Crop Breeding. A.C.; Fasoulas, P.O. Box 1555, GR, 54006.Thssaloniki 17, Greece. 1993.;
8. Wuhaib, K.M.Testing introduced maize germplasm by line´tester method I- yield and yield components. The Iraqi J. of Agric Sci, 2012.; 43(1): 38-48.
9. Wuhaib, K.M.; Hadi, B.H.; Hassan, W.A. Hybrid vigor, heterosis, and Genetic parameters in maize by diallel cross analysis. Int. J. of Applied Agric. Sci. 2016.; 2(1): 1-11.
10. Dubey, R.B.; Joshi, V.N.; Pandiya, N.K. Heterosis and combining ability for quality, yield maturity traits in conventional and nonconventional hybrids of maize (Zea mays L.) Ind. J. Genet. Plant Breed. 2001.; 61:353-355.
11. Abed, N.Y.; Hadi, B.H.; Hassan, W.A.; Wuhaib, K.M. Assessment yield and its components of Italian maize inbred lines by full diallel cross. Al-Anbar J. of Agric, Sci. .2017.;15:114-124.
12. Wannows, A.A.; Azzam, H.K.; AL-Ahmad, S.A. Genetic variances, heritability, correlation and path coefficient analysis in yellow maize crosses (Zea mays L.). Agric. Biol. J. N.Am. 2010.; 1(4): 630-637.
13. Singh, R.K.; Chaudhary, B.D. Biometrical Methods in Quantitative Genetic Analysis. Kaly-ani Publishers, New Delhi. 2007.;.
14. Mezmouk, S.; Mumm, R.H.; Ross-Ibarra, J. Heterosis and genetic load investigated in partial diallel cross. Plant and Animal Genome XXII Conference (USA). 2014.; P. 178.
15. Alaminie, A.; Wali, M.C.; Salimath, P.M.; Jagadeesha, R.C. combining ability and heterosis for grain yield and ear characters in maize. Karnataka J.Agric. Sci. 2006.; 19:13-16.
16. Nigussie, M.; Zelleke, H. Heterosis and combining ability in diallel among eight elite maize population. African Crop Sci. 2001.; J. 9(3): 471-479.
17. Srdic, J.; Drinic, S.M.; Pajic, Z.; Filipovic. M. Characterization of maize inbred lines based on molecular markers, heterosis and pedigree data. Genetika, 2007.;39(3): 355-363.
18. Aliu, S.; Fetahu, S.; Salillari, A. Estimation of heterosis and combining ability in maize (Zea mays L.) for ear weight using diallel crossing method. Latvian J. of Agron. 2008.; 11: 7-11.
19. Barznji, M.O.H. Estimation of some genetic parameters using full diallel cross in maize. Thesis. Field Crop. Univ. of Sulaimania. 2013.; Pp: 98.
20. Iqbal, A.M.; Nehvi, F.A.; Wani, S.A.; adir, R.Q.; Dark, Z.A. Combining ability analysis for yield and yield related traits in maize (Zea mays L.). Int. J. Plant Breed. Genet. 2007.; 1: 101-105.
21. Abedalhammed, H. S., Naser, A. S., Al-Maathedy, M. H., Mohammed, Th. T., Jaber, B. T. & Al-Asha'ab, M. H. The effect of vitamin e as an antioxidant with different levels of dried tomato pomace supplementation on diets of common carp (cyprinus carpio l) on blood indices. Biochemical and Cellular Archives. 2020, 20(2): 5173-5176. .
22. Dadheech, A.; Joshi, V.N. Heterosis and combining ability for quality and yield in early maturing single cross hybrids of maize (Zea mays L.). Indian J. Agric. Res. 2007.; 41:210-214.
23. Barakat, A.A.; Osman, M.M.A. Evaluation of some newly developed yellow maize inbred lines for combining ability in two locations. J. Agric. Sci. 2008.; 33: 4667-4679.
24. F. T. Al-Rawi, Y. T. Abdul-Rahaman , Abdullah I.Noaman , Th. T. Mohammed, S. M Abdulateef, Nadia Jebril and K.I. Mahmud. Role of ascorbic acid and appetite stimulants on a few blood serum biochemical characteristics in pregnant Iraqi ewes under heat stress. Al-Rawi F T, Abdul-Rahaman Y T, Noaman Revis Bionatura 2022;7(4) 6. http://dx.doi.org/10.21931/RB/2022.07.04.6.
25. Z. Al-Fayyadh, D. .; Hasson, A. A. .; Hussein, A. K. .; Hassan, R. K. Effect Of Humic Acid Spray On Growth Characteristics Of Wheat Varieties . Journal of Life Science and Applied Research. 2020, 1, 10-19.
Received: January 15, 2023 / Accepted: February 25, 2023 / Published:15 March 2023
Citation. Hadi, B.H.; Hassan, W.A.; Alshugeairy. Z.K,; Alogaidi. F.F. Estimating genetic parameters of maize hybrids and parents under different plant densities (Combining ability for yield and some other traits for maize Zea mays L.). Revis Bionatura 2023;8 (1) 87 http://dx.doi.org/10.21931/RB/2023.08.01.87