2017.02.02.8
Files > Volume 2 > Vol 2 No 2 2017 > Revisiones
ARTÍCULO DE REVISIÓN
Estrategias Naturales para Mejorar el Crecimiento y la Salud en los Cultivos Masivas de Camarón en Ecuador
Natural strategies to Improve Growth and Health in Massive Shrimp Cultures in Ecuador

Luis E. Trujillo1*, Leonor Rivera2,3, Eugenio Hardy1,4, Erika M. Llumiquinga1, Francisco Garrido1, Jesús A. Chávez3, Victor H. Abril1, José M. País-Chanfrau5
RESUMEN
El camarón, es actualmente el segundo producto comercial no petrolero más grande de Ecuador, debe cultivarse preferentemente con productos orgánicos naturales que no sean productos químicos sintéticos o antibióticos. Por lo tanto, los probióticos, los prebióticos y los inmunoestimuladores tales como los lipopolisacáridos (LPSs) actualmente atraen gran atención para el cultivo de camarón, la gestión de la calidad del agua y el tratamiento de enfermedades acuáticas. De hecho, en diferentes países, los probióticos basados en bacterias, levaduras o sus componentes se han utilizado con éxito en esta práctica. También se sugiere ampliamente el uso de prebióticos como aditivos alimentarios para camarones. Por otra parte, se conoce que los lipopolisacáridos, que son el componente principal de la membrana externa de las bacterias gramnegativas, ejercen fuertes efectos sobre el sistema inmune del camarón. La presente revisión destaca el uso de estas importantes entidades biológicas como potenciales candidatos a ser utilizados en la acuicultura ecuatoriana. A pesar de que una gran variedad de efectos positivos de los probióticos, prebióticos y LPSs en peces y crustáceos han sido descubiertos, aún se necesita más información para aclarar y prever el impacto molecular de estas entidades biológicas en camarones.
Palabras claves: Prebióticos, probióticos, fructoligosacáridos, lipopolisacáridos, camarón, levadura.
_______________________________________________________________________________________________________________________
ABSTRACT
Shrimp, which is currently the second largest non-oil commercial product from Ecuador, should be preferably cultivated with natural organic products other than synthetic chemicals or antibiotics. Therefore, probiotics, prebiotics and immunostimulators such as lipopolysaccharides currently attract great attention for shrimp culturing, water quality management and treating aquatic diseases. In fact, probiotics based on either bacterium, yeasts or its components have been used successfully in practice. Also, the use of prebiotics as food additives for shrimps is extensively suggested. Lipopolysaccharides, which are the major component of the outer membrane of gram-negative bacteria, are known to exert strong effects on the shrimp immune system. The present review highlights the use of these important biological entities as potential candidates to be used in Ecuadorian shrimp aquaculture. Despite a variety of positive effects of probiotic, prebiotic and LPS supplements on fish and crustaceans have been uncovered, significantly more information is still required in order to clarify and foresee the molecular impact of these biological entities in shrimps.
Key words: Prebiotic, probiotic, fructoligosaccharides, lipopolysaccharide, shrimp, yeasts.
_______________________________________________________________________________________________________________________
INTRODUCCIÓN
A finales de 1996, Ecuador se convirtió en el segundo mayor exportador mundial de camarones1. En la actualidad, el camarón es el segundo producto no petrolero que comercialmente más aporta a la economía después de los bananos en Ecuador. Durante 2013, por ejemplo, 474 236 libras de camarón fueron exportadas fuera de Ecuador, lo que equivale a 1 620 millones de dólares. En 2014, se exportaron medio millón de libras de camarón, generando un ingreso de 1 520 millones de dólares. La producción de camarón sigue creciendo de manera constante como se muestra en la figura 1, por lo que se espera que sus exportaciones superen los valores previos a fines de este año. Cabe destacar que Ecuador sigue siendo el mayor productor latinoamericano y el quinto productor mundial de camarones de cultivo2. En consecuencia, la cría de camarón conduce a un mayor intercambio de divisas y una mayor fuente de mano de obra en este país.
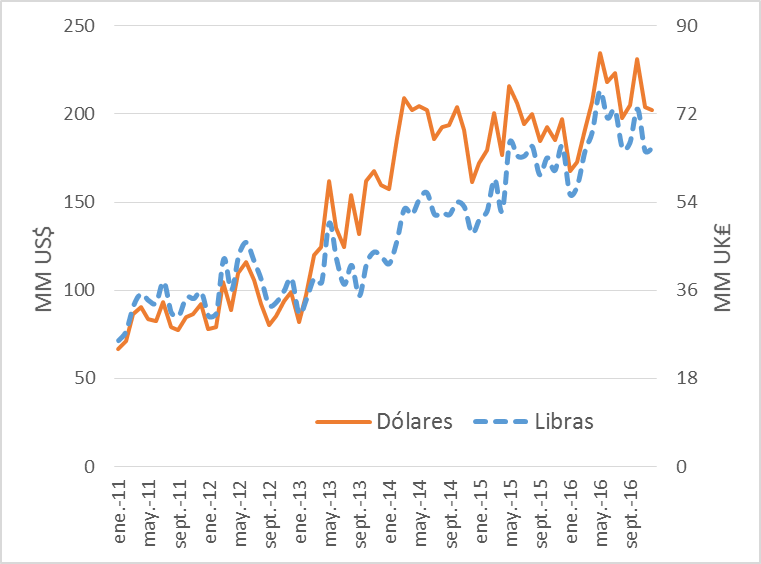
Figura 1. Exportación de camarones hasta finales de 2016 Vs Millones de dólares obtenidos 2.
La producción masiva de camarones no está exenta de riesgos. De hecho, la aparición del virus de mancha blanca y otros brotes de enfermedades en el siglo pasado han causado una disminución en la producción de camarón, afectando así social y económicamente al país3. Otros problemas recientes asociados con el cultivo intensivo de camarón son una disminución del agua4 y la calidad de los alimentos, un aumento del estrés animal y un aumento de infecciones bacterianas, virales o parasitarias.
Los antibióticos se han utilizado tradicionalmente para tratar las infecciones microbianas en la acuicultura en todo el mundo. Sin embargo, el uso y el abuso de antibióticos puede causar efectos adversos no deseados como la acumulación de antibióticos en los tejidos de los animales, supresión inmune, desarrollo de bacterias resistentes a los antibióticos y destrucción de la flora microbiana ambiental5,6. Por otro lado, el uso de antibióticos para el tratamiento de peces y camarones es costoso y, por consiguiente, no está disponible en muchas granjas ecuatorianas. Por lo tanto, las investigaciones encaminadas a la sustitución de antibióticos por productos naturales como los probióticos, los prebióticos y los lipopolisacáridos (LPS) han recibido recientemente una atención considerable por parte de los investigadores del camarón y de estos productores naturales alternativos en Ecuador. Este hecho asegura el desarrollo de diferentes bioprocesos para obtener estas moléculas en el país7,8.
El término probiótico significa "para la vida", y se deriva de las palabras griegas "pro" y "bios"9. La definición más amplia de probióticos fue formulada por Fuller10 indicando que un probiótico es considerado como "un suplemento alimenticio microbiano vivo que afecta beneficiosamente al animal huésped mejorando su equilibrio intestinal". Sin embargo, no sólo los microorganismos vivos deben ser considerados como probióticos, sino también cualquier microorganismo o cualquiera de sus componentes que puedan conferir cualquier efecto fisiológico beneficioso al huésped o al medio ambiente donde vive el microorganismo.
Un prebiótico, a diferencia de un probiótico, no es un organismo y por lo tanto tiene menos influencia en el medio ambiente natural. Basándose en la definición de Gibson y Roberfroid, los prebióticos son ingredientes alimentarios no digeribles que afectan de manera beneficiosa al huésped al estimular selectivamente el crecimiento y / o la actividad de uno o un número limitado de bacterias en el colon y así mejorar la salud del huésped11,12. Los prebióticos tienen dos características principales, la primera es que son alimentos naturales, y la segunda es que no son organismos vivos, por lo que este hecho constituye una ventaja sobre los probióticos.
La incorporación de prebióticos en la dieta no necesita autorización como aditivos alimentarios ya que están bien descritos en la literatura. Originalmente, los prebióticos se utilizaron para estimular bifidobacterias y lactobacilos en la microbiota humana13.
Los lipopolisacáridos inducen efectos fuertes en el sistema inmune del camarón. Estos efectos han sido también objeto de numerosos artículos (véase más adelante) en los que se presentan casos interesantes como evidencia de la posible aplicación de LPS como inmunoestimulante en la cría de camarón.
La presente revisión destaca el uso de probióticos, prebióticos y LPS, mecanismos de acción y retos futuros en el uso y producción de estas moléculas en la acuicultura ecuatoriana a base de experiencias nacionales e internacionales.
LOS PROBIÓTICOS EN LA ACUICULTURA
Los probióticos son formulaciones microbianas cultivadas que se mezclan con enzimas y cereales fermentados unidos a un sustrato o formulación de uso en acuicultura14. En el campo de los organismos acuáticos, los probióticos se han definido como "células microbianas capaces de viajar a través del tracto gastrointestinal y de mantenerse vivas, con el objetivo de mejorar la salud"15,16. Una definición más amplia postula que "los probióticos se definen como entes vivos o muertos, o incluso un componente de los microorganismos que actúan bajo diferentes modos de acción al conferir efectos beneficiosos no sólo al huésped sino también a su medio ambiente”.
Existe un interés creciente por los probióticos17 que ha sido estimulado por varios factores, entre ellos: 1) resultados científicos y clínicos interesantes utilizando organismos probióticos bien documentados; 2) inquietudes sobre efectos secundarios y daños ambientales; Para el uso de productos naturales, y (4) el desarrollo de interesantes antagonistas acuáticos para la acuicultura ecológica. La Tabla 1 muestra cepas microbianas probadas con éxito como probióticos en condiciones de campo.

Tabla 1. Cepas microbianas autorizadas y probadas utilizadas como probióticos en condiciones de campo.
Los probióticos funcionan de acuerdo con mecanismos de exclusión competitiva por los cuales los patógenos son reemplazados o excluidos por el desarrollo de una población microbiana beneficiosa en la superficie intestinal, lo que reduce las incidencias de la enfermedad, mejor salud y mejor crecimiento del huésped (Fig.2).
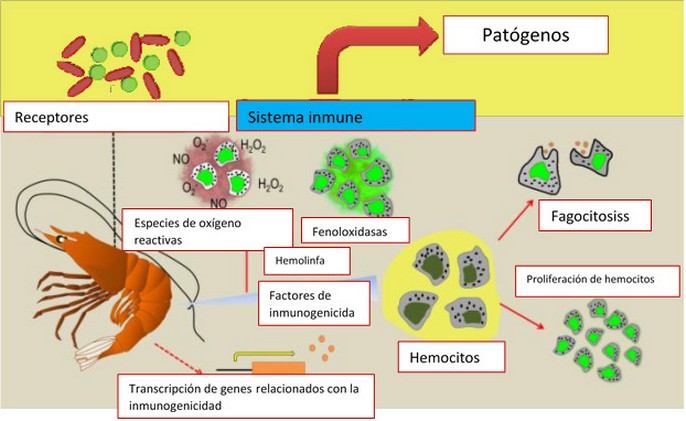
Figura 2. Como actúan los probióticos en el camarón83. La inhibición directa de patógenos mediante la modulación del sistema inmune del camarón, la producción de sustancias inhibidoras que podrían interactuar directamente con patógenos y también son capaces de manipular la microflora y las condiciones físico-químicas del ecosistema del estanque.
Las cepas bacterianas, de levadura o sus componentes son capaces de producir sustancias inhibidoras que podrían interactuar directamente con patógenos bacterianos y virales. El crecimiento de estos patógenos se evita a través de la producción de metabolitos antagonistas24–26 o por interferencia de adhesión27.
Algunos de los estudios previos realizados en los primeros 90 años del siglo pasado, reportados por Nogami y Maeda28, y Rengpipat et al.29 mostraron un mejor crecimiento y supervivencia de crustáceos (camarones, cangrejos) con suplementos probióticos.
Algunos informes internacionales y nacionales describen un aumento significativo en el peso de camarones tratados con probióticos sobre el grupo no tratado a nivel de finca30 como se muestra en la Tabla 2.
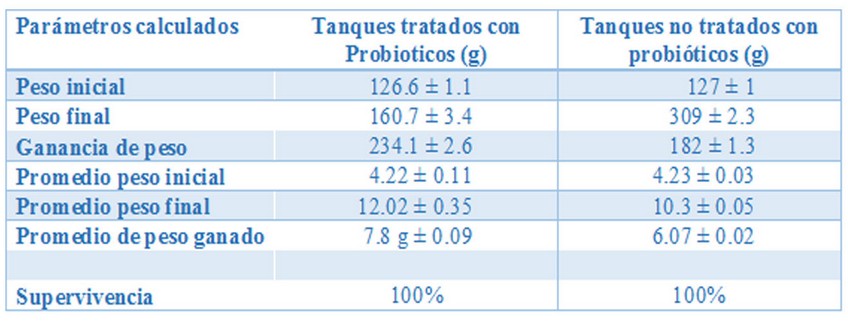
Table 2. Datos experimentales obtenidos en tanques de cultivo de camaron P. monodon tratados y no tratados con probóticos.
Lara-Flores et al.31 explicó que las bacterias probióticas son buenos candidatos para aumentar la digestión de nutrientes y el crecimiento en organismos acuáticos. Irianto y Austin32 afirmaron que los compuestos nocivos en la dieta pueden ser destoxificados por la acción de enzimas hidrolíticas tales como amilasas y proteasas, y también que los nutrientes en los organismos podrían ser mejorados por vitaminas como por ejemplo, biotina y vitamina B12. Las enzimas digestivas son necesarias para romper los compuestos complejos en moléculas más simples y absorbibles que podrían ser utilizadas por el huésped33. Por lo tanto, la suplementación dietética con probióticos aumenta el crecimiento34 a través de la contribución enzimática que aumenta la digestibilidad35. Los constituyentes de la digestión de proteínas generados por la acción de enzimas proteolíticas endógenas también contribuyen a aumentar la digestibilidad de los piensos asociados con la administración de probióticos35
.
Se han propuesto probióticos para mejorar la respuesta inmune de los camarones. De hecho, la bacteria probiótica Lactobacillus plantarum mejora la expresión génica y las respuestas inmunes en el camarón blanco Litopenaeus vannamei, cuando se combina en su dieta. Específicamente, L. plantarum mejora la actividad de la fenoloxidasa, la profenoloxidasa (proPO) y la superóxido dismutasa (SOD), junto con la tasa de supervivencia después de la estimulación con V. alginolyticus. Estos efectos se observan cuando las bacterias se dan en la dieta a 1010 ufc/kg durante 168 horas36
L. plantarum tiene una gran efectividad contra Vibrio harveyi. Desafíos experimentales in vivo, muestra que Litopenaeus vannamei aumentó la resistencia en comparación con un grupo control no expuesto a V. harveyi. Estos resultados han sugerido que la cepa probiótica tiene un efecto inmuno-reactivo sobre el huésped, posiblemente relacionado con compuestos extracelulares liberados de bacterias que pueden estimular respuestas inmunes inespecíficas. La actividad de L. plantarum es responsable del aumento del recuento total de hemocitos y de la actividad PO. De acuerdo con los resultados de esta investigación, se sugirió que la actividad aglutinante es la causa principal relacionada con la eliminación más efectiva de la hemolinfa y hepatopancreas por el camarón alimentado con la dieta suplementada con probiótico37.
Cuando se desafió con Vibrio nigripulchritudo, el probiótico Pediococcus acidilactici influyó en las defensas antioxidantes y el estrés oxidativo de Litopenaeus vannamei38. Este probiótico muestra eficacia en marcadores de defensa antioxidantes como SOD, catalasa, glutatión peroxidasa, glutatión y daño tisular. Esta cepa fue eficiente en el mantenimiento de los niveles de defensa antioxidante durante un período más largo que el control y los grupos de camarones no infectados39.
Entre las tres cepas, Bacillus P64 muestra un índice de inmunidad significativamente mayor y una respuesta inmune similar a la de V. alginolyticus. Además, el Bacillus P64 induce un aumento en la población de células hialinas40,41.
Algunos otros Vibrio sp. que han sido probados por su actividad probiótica en L. vannamei son V. alginolyticus (NCIMB 1339) y V. gazogenes (NCIMB 2250). Estas cepas muestran actividad antagonista contra los patógenos del camarón. Cuando los camarones juveniles son alimentados con quitina y V. gazogenes, el número de bacterias tipo vibrio en la tripa anterior y posterior disminuye significativamente y el número de hemocitos cambia significativamente. El cambio en el número de hemocitos probablemente refleja el estado inmunológico del camarón debido a que los hemocitos están involucrados tanto en defensas celulares como humorales del camarón42.
Los β-glucanos de levadura también se han aplicado como probióticos en la acuicultura para modular el sistema inmune innato de los peces y para mejorar la supervivencia de los peces frente a patógenos43,44. Si se administran como aditivos alimentarios, los β-glucanos pueden ejercer sus efectos primarios a través de la inducción de citoquinas, que a su vez afectan a la respuesta inmune sistémica en los peces. Se han evaluado diferentes fuentes de β-glucanos, aunque las fuentes más frecuentes son la levadura panadera, Saccharomyces cerevisiae. Algunas preparaciones comerciales de β-glucano que se pueden usar en acuicultura son MacroGard®, Betagard A®, EcoActivaTM, NutrifermTM, BG y Fibosel®). Muchos estudios han explorado la respuesta in vitro de los macrófagos al β-glucano45 mientras que otros estudios han abordado el efecto in vivo de β-glucanos en diferentes especies de peces46. Los β-glucanos de varias fuentes se han administrado a los peces por vía oral o intraperitoneal con diferentes efectos sobre la producción de anticuerpos, la expresión de genes del sistema inmune, la supervivencia, la resistencia a enfermedades infecciosas y la mejora en la resistencia al estrés. Se ha informado que algunos productos probióticos como Superbióticos, SuperPs, Zymetin y Mutagen desempeñan un papel vital en postlarvas de P. monodon manteniendo buenos parámetros de calidad del agua a lo largo del período de cultivo47. Pais-Chanfrau et al. 201648 modificaron el medio de cultivo para obtener grandes cantidades de biomasa de levadura para diferentes aplicaciones incluyendo la producción de β-glucanos para diferentes aplicaciones en cultivos masivos de camarón.
LOS PREBIÓTICOS EN LA ACUICULTURA
Los prebióticos, a diferencia de los probióticos, no son organismos vivos por lo que tienen menos influencia en el medio ambiente. Mahious et al.49, Fooks et al.50 y Gibson et al.12 describieron que cualquier alimento que llega al colon por ejemplo, carbohidratos no digeribles, péptidos, proteínas y ciertos lípidos, puede ser un candidato a prebiótico. Sin embargo, los carbohidratos no digeribles como oligosacáridos, inulina, transgalactooligosacárido y lactulosa son considerados como el foco de la mayoría de los oligosacáridos12. En peces, los oligosacáridos de manano (MOS ) y FOS son quizás los oligosacáridos más bien estudiados a pesar de la información sobre el uso de otros compuestos similares a oligosacáridos en animales acuáticos que también está disponible en la literatura51,52. La estructura química de algunos de estos oligosacáridos se presenta en la figura 2.
Bailey et al.53 y otros autores54,55 informaron que los prebióticos pueden modificar la comunidad microbiana del tracto gastrointestinal para potenciar las respuestas inmunes no específicas. La fermentación prebiótica en el colon por las bacterias causa una modificación significativa de la microflora colónica. De hecho, los oligosacáridos sirven como sustrato para el crecimiento y la proliferación de bacterias anaerobias, principalmente las bifidobacterias, que inhiben el crecimiento de bacterias putrefactivas y patógenas presentes en el colon56.
La producción resultante de sustancias que estimulan el sistema inmunológico aumenta la protección del huésped contra las infecciones. Li et al.57 encontraron que la suplementación con FOS de cadena corta a concentraciones de 0,025 a 0,8% (m/m) aumenta la capacidad respiratoria de hemocitos en el camarón blanco del Pacífico Litopenaeus vannamei cultivado en un sistema con recirculación. La suplementación dietética de Grobiotic®-A mejora la supervivencia del camarón blanco del pacífico cultivado en agua de baja salinidad. Además, un desafío de agua dulce también muestra una mejora similar en la supervivencia del camarón alimentado con Grobiotic®-A, aunque no se han identificado los mecanismos para mejorar la supervivencia bajo condiciones de baja salinidad58.
Los prebióticos pueden modificar la comunidad microbiana del tracto gastrointestinal para mejorar las respuestas inmunes no específicas53. También ayudan a reducir el pH del intestino a través de la formación de ácidos grasos de cadena corta, lo que favorece las bacterias beneficiosas que bloquean la adhesión de los patógenos y los sitios de los receptores de toxinas, la competición de nutrientes y la estimulación del sistema inmunológico50,59,60.
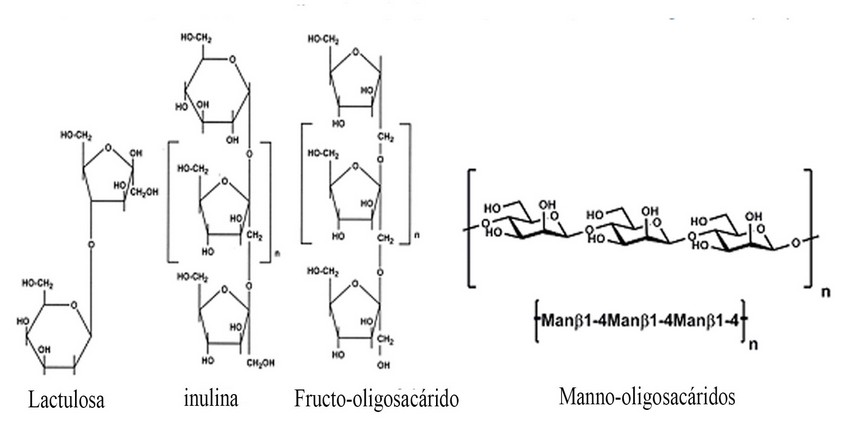
Figura 3. Estructura quimica de los fructo-oligosacáridos (FOS), la inulina, la lactulose y los manno-oligosacaridos (MOS).
Sin embargo, los informes sobre el efecto de los prebióticos sobre los parámetros de crecimiento en los peces no son concluyentes y algunos son contradictorios. La dieta de Beluga (Huso huso) suplementada con 1, 2 y 3% (m/m) de inulina ha afectado negativamente a una variedad de parámetros tales como aumento de peso, crecimiento, relación de eficiencia proteica, proteína y retención de energía, eficiencia alimentaria al suplementar con inulina. Además, los parámetros de crecimiento en los peces alimentados con inulina son inferiores a los mostrados en el grupo de control. La reducción de algunos de estos parámetros, especialmente el crecimiento, en los grupos tratados puede deberse al efecto de parámetros distintos de los relacionados con la inulina61. En cualquier caso, la inulina no ha mostrado efectos positivos en la tasa de crecimiento de los peces jóvenes Huso huso.
La dieta de salmón del Atlántico suplementada con 10 g/kg de prebióticos como MOS, FOS y galacto-oligosacárido (GOS) no muestró efectos sobre el crecimiento y la digestibilidad62. La eficacia de la alimentación mejora significativamente cuando se utiliza una dieta suplementada con 10-20 g/kg de Grobiotic®-AE durante 7 semanas, pero el crecimiento no es significativo63. Además, Refstie et al.64 encontraron que el salmón atlántico alimentado durante 3 semanas con una dieta basada en harina de pescado suplementada con 75 g/kg de inulina aumenta la masa relativa del tracto gastrointestinal, pero la capacidad de absorción del pez no se ve afectada. Una dieta con 20 g/kg de oligofructosa, un FOS producido por hidrólisis enzimática parcial de inulina, aumenta el crecimiento de larvas de rodaballo, pero 20 g/kg de inulina en sí misma no tiene efecto sobre el crecimiento65. El crecimiento, la eficiencia de la alimentación y la supervivencia se han mejorado en dos experimentos con trucha arco iris alimentada con una dieta que contiene 2 g/kg de MOS comparado con las truchas alimentadas con dieta basal62.
La concentración de proteína corporal en la trucha arco iris y la tilapia híbrida aumenta cuando el nivel de MOS aumenta en la dieta de 1,5 a 4,5 g/kg66. Por otra parte, una dieta suplementada con 10 g/kg de MOS o GOS, resulta en una disminución de la concentración de proteína en el cuerpo del salmón, lo cual está en línea con los resultados de Genc et al.67 que alimentaron a Penaeus semisulcatus con una dieta que contenía 4,5 g/kg de MOS. Estos autores especularon que la concentración de proteínas en el cuerpo inferior en camarones alimentados con la dieta MOS puede haber sido el resultado de una menor utilización de aminoácidos y la digestibilidad de la dieta62. La suplementación dietética con inulina no afecta la composición corporal de la Beluga juvenil68. A pesar de estos resultados contradictorios, la ingesta de prebióticos depende del tipo de ingredientes utilizados en las formulaciones de la dieta y, por lo tanto, variará entre las especies y las dietas. El tipo de prebiótico para suplementar, características animales específicas (especie, edad, etapa de producción) y tipo de dieta se han identificado como consideraciones importantes al suplementar prebióticos en dietas de pescado. Sin embargo, muchos resultados experimentales no han sido concluyentes con respecto al beneficio de algunos de estos prebióticos. No obstante, debe prestarse atención al desarrollo de bioprocesos, formulaciones prácticas y consideraciones económicas que aseguren la producción rentable y la disponibilidad de FOS y GOS para diferentes aplicaciones en el campo de los organismos acuáticos y la alimentación animal en general7.
LOS LIPOPOLISACÁRIDOS COMO INMUNOESTIMULADORES
Los efectos del LPS sobre el sistema inmunológico del camarón han sido objeto de numerosos artículos. Los casos interesantes que pueden aportar evidencia de la posible aplicación de LPS como inmunoestimulante en la cría de camarón se describen a continuación. Los porcentajes de proliferación de hemocitos pueden aumentar, aproximadamente tres veces, después de la estimulación con LPS. Además, la captación de 3H timidina en los hemocitos circulantes puede ser significativamente 26 veces mayor en camarones estimulados con LPS que en los no estimulados69.
El sistema de activación proPO del Penaeus paulensis mejora en gran medida con el tratamiento con LPS, lo que sugiere su participación en el no auto-reconocimiento70. Además, la actividad PO en el suero de camarón aumentó con el tratamiento con LPS. La administración oral de Pantoea agglomerans con LPS mejora la resistencia a la enfermedad frente a la viremia aguda de los peneidos e induce la actividad inactivadora de los virus en la hemolinfa del camarón Kuruma, Penaeus japonicas71. La proliferación de la mayoría de las células en el tejido hematopoyético del camarón tigre negro (Penaeus monodon) puede incrementarse significativamente después de la inyección de LPS72. La inyección de camarón blanco del Pacífico Litopenaeus vannamei con LPS disminuyó los niveles de ARNm de péptidos antimicrobianos, penaeidin 2, penaeidin 3, penaeidin 4 de una manera dependiente de la dosis, mientras que los niveles de ARNm de serin proteinasa y proPO no cambian significativamente73. La estimulación de los hemocitos del camarón Litopenaeus vannamei con LPS conduce a la formación de trampas extracelulares características de fibras hechas a partir del ADN de la célula huésped74. Los grupos de camarón tigre negro (Penaeus monodon) alimentados con gránulos recubiertos con LPS mostraron mayores tasas de supervivencia que los grupos de control cuando se desafíaron frente al Vibrio harveyi 75.
Además, los niveles de genes relacionados con la inmunidad com por ejemplo, la isoforma 3 del factor anti-LPS, la lectina de tipo C y la peritrofina de tipo mucina, están regulados en los intestinos del grupo de P. monodon alimentado con dieta suplementada con LPS una o dos veces al día, a diferencia de del grupo de control. La expresión de lectina 3 de tipo C de Litopenaeus vannamei en branquias está regulada positivamente después del desafío contra LPS76. Los LPSs pueden regular la cactina de L. vannamei en hemocitos77. La incubación de hemocitos de camarón con LPS puede causar necrosis y liberar moléculas endógenas como por ejemplo, L, β y P, mientras que al mismo tiempo estas moléculas endógenas pueden causar cambios en la viabilidad celular, desgranulación y necrosis de hemocitos, indicativo de activación de la inmunidad in vitro78. La inyección de LPS causa una disminución de los recuentos totales de hemocitos semi-granulares y granulares de Penaeus monodon79. Además, la actividad de esterasa no específica, la producción de especies reactivas de oxígeno y la producción de óxido nítrico se induce significativamente en los hemocitos de camarón inyectado con LPS, mientras que la relación de células apoptóticas de hemocitos aumenta. Recientemente se han informado métodos para aislar y caracterizar los LPS con el fin de usarlos en el cultivo del camarón8.
OBSERVACIONES FINALES
Ecuador y la India son actualmente los dos países que lideran la exportación de camarón (http://www.fao.org/in-action/globefish/market-reports/resourcedetail/en/c/462149/). Los tres princi-pales destinos de exportación hasta 2016 de la industria del camarón ecuatoriano fueron Estados Unidos (35 000 toneladas), Unión Europea (44 000 toneladas) y Vietnam (80 000 toneladas). Por lo tanto, el diseño de bioprocesos para obtener un conjunto de moléculas para mejorar las técnicas biotecnológicas es necesario para hacer frente al aumento de la producción de camarón, contribuyendo así al éxito de esta industria.
La aplicación de probióticos, prebióticos80 y LPS en acuicultura ha mostrado resultados positivos, pero la evaluación de los efectos biológicos de estas entidades en el medio natural y la rentabilidad de las opciones de tratamiento son insuficientes. Además, la mezcla de estos reactivos biológicos para formar simbióticos tiene un campo muy prometedor en cultivos de camarón81 ya que esta estrategia ha tenido cierto éxito en otras especies marinas82.
Siguiendo las numerosas herramientas de secuenciación del genoma que están actualmente en uso, la investigación futura sobre probióticos, prebióticos y efectos de LPS debería involucrar el análisis de transcriptome y proteoma usando ensayos de alto rendimiento. Además, se debe estudiar el desarrollo de transcriptomas y proteomas en la microbiota intestinal. Desde el punto de vista bacteriológico, es de gran importancia continuar evaluando si los cambios microbianos tienen algún efecto positivo sobre la salud de los peces y de las conchas, contribuyen a inhibir la colonización de las bacterias patógenas en el intestino y mejorar la inmunidad innata. Por lo tanto, los estudios con todas estas sustancias deberían tener una alta prioridad en el futuro, y los estudios de desafíos deberían incluirse como un estándar de oro para evaluar sus efectos sobre la salud del camarón.
REFERENCIAS
1. Chavez H, Herrera Z. Análisis Estadítico de la Producción Camaronera del Ecuador. 2012. https://www.dspace.espol.edu.ec/bitstream/ 123456789/2139/1/ 4218.pdf.
2. Wurmann G. C. REGIONAL REVIEW ON STATUS AND TRENDS IN AQUACULTURE DEVELOPMENT IN LATIN AMERICA AND THE CARIBBEAN – 2015.; 2017. http://www.fao.org/3/a-i6867e.pdf.
3. Lescano E. Implementacion de la nic 41 en un empresa camaronera. 2011. http://repositorio.ucsg.edu.ec/bitstream/3317/1409/1/T-UCSG-PRE-ECO-CICA-16.pdf.
4. Ghosh AK, Bir J, Azad MAK, Hasanuzzaman AFM, Islam MS, Huq KA. Impact of commercial probiotics application on growth and production of giant fresh water prawn (Macrobrachium Rosenbergii De Man, 1879). Aquac Reports. 2016;4:112-117. doi:10.1016/j.aqrep.2016.08.001.
5. Romero J, Feijoó CG, Navarrete P. Antibiotics in aquaculture – Use, abuse and alternatives. In: Health and Environment in Aquaculture. ; 2012:160-198. doi:10.5772/28157.
6. Hernández Serrano P. Responsible use of antibiotics in aquaculture. FAO Fish Tech Pap. 2005;469(November 2011):97 pp. doi:10.1007/s13398-014-0173-7.2.
7. Trujillo LE, VE. Marcillo, R. Avalos, LK. Ponce TR. From the laboratory to the industry: Enzymatic production and applications of short chain fructoligosaccharides (FOS): Recent advances and current perspective. Bionatura. 2015;1(1):12-16. http://revistabionatura.com/index.php/Bionatura/article/view/16.
8. Hardy E, Rodriguez C, Trujillo LE. Lipopolysaccharide (LPS) and protein-LPS complexes: Detection and characterization by gel electrophoresis, mass spectrometry and bioassays. Biol Med. 2016;8(3). doi:10.4172/0974-8369.1000277.
9. Gismondo MR, Drago L, Lombardi A. Review of probiotics available to modify gastrointestinal flora. Int J Antimicrob Agents. 1999;12(4):287-292. doi:10.1016/S0924-8579(99)00050-3.
10. Fuller R. Probiotics in man and animals. J Appl Bacteriol. 1989;66(5):365-378. doi:10.1111/j.1365-2672.1989.tb05105.x.
11. Gibson GR, Roberfroid MB. Dietary Modulation of the Human Colonic Microbiota: Introducing the Concepts of Prebiotics. J Nutr. 1995;125(6):1401-1412. isi:A1995RC95800001.
12. Gibson GR, Probert HM, Loo J Van, Rastall RA, Roberfroid MB. Dietary modulation of the human colonic microbiota: updating the concept of prebiotics. Nutr Res Rev. 2004;17:259-275. doi:10.1079/NRR200479.
13. Gatesoupe F. Probiotics and prebiotics for fish culture , at the parting of the ways. Aqua Feed Formul Beyond. 2005;2(3):3-5. http://www.feedware.com/index.html.
14. Ravichandran S, Rameshkumar G, Prince AR. Biochemical Composition of Shell and Flesh of the Indian White Shrimp Penaeus indicus (H.milne Edwards 1837). Am J Sci Res. 2009;4(3):191-194.
15. Gatesoupe FJ. Probiotic and formaldehyde treatments of Artemia nauplii as food for larval pollack, Pollachius pollachius. Aquaculture. 2002;212(1-4):347-360. doi:10.1016/S0044-8486(02)00138-2.
16. Lazado CC, Caipang CMA. Atlantic cod in the dynamic probiotics research in aquaculture. Aquaculture. 2014;424-425:53-62. doi:10.1016/j.aquaculture.2013.12.040.
17. Kumar A. Probiotics: An effective feed supplement in shrimp culture. J Bangladesh Acad Sci. 2014;38(2):127-130.
18. Moreira D, Sabrina M, Leivas FP, Romano LA, Ballester E. New Bacillus Probiotic Tested for Shrimp. Glob Aquac Advocate. 2011;Jan/Feb:46-47.
19. Moriarty DJW, Decamp OP, Lavens P. Probiotics in Aquaculture. AQUA Cult Asia Pacific Mag. 2005;1:14-16.
20. Utiswannakul P, Sangchai S, Rengpipat S. Enhanced growth of black tiger shrimp Penaeus monodon by dietary supplementation with Bacillus (BP11) as a probiotic. J Aquat Res Dev. 2011;3:2155–2164.
21. Scholz U, Garcia Diaz G, Ricque D, Cruz Suarez LE, Vargas Albores F, Latchford J. Enhancement of vibriosis resistance in juvenile Penaeus vannamei by supplementation of diets with different yeast products. Aquaculture. 1999;176(3-4):271-283. doi:10.1016/S0044-8486(99)00030-7.
22. Ajitha S, Sridhar M, Sridhar N, Singh ISB, Varghese V. Probiotic effects of lactic acid bacteria against Vibrio alginolyticus in Penaeus (Fenneropenaeus) indicus. Asian J Fish Sci. 2004;17:71–80.
23. Villamil L, Figueras A, Planas M, Novoa B. Control of Vibrio alginolyticus in Artemia culture by treatment with bacterial probiotics. Aquaculture. 2003;219(1-4):43-56. doi:10.1016/S0044-8486(02)00515-X.
24. Chythanya R, Karunasagar I, Karunasagar I. Inhibition of shrimp pathogenic vibrios by a marine Pseudomonas I-2 strain. Aquaculture. 2002;208(1-2):1-10. doi:10.1016/S0044-8486(01)00714-1.
25. Iyapparaj P, Maruthiah T, Ramasubburayan R, et al. Optimization of bacteriocin production by Lactobacillus sp. MSU3IR against shrimp bacterial pathogens. Aquat Biosyst. 2013;9(1):12. doi:10.1186/2046-9063-9-12.
26. Lazado CC, Caipang CMA, Rajan B, Brinchmann MF, Kiron V. Characterization of GP21 and GP12: Two potential probiotic bacteria isolated from the gastrointestinal tract of Atlantic cod. Probiotics Antimicrob Proteins. 2010;2(2):126-134. doi:10.1007/s12602-010-9041-8.
27. Lazado CC, Caipang CMA, Brinchmann MF, Kiron V. In vitro adherence of two candidate probiotics from Atlantic cod and their interference with the adhesion of two pathogenic bacteria. Vet Microbiol. 2011;148(2-4):252-259. doi:10.1016/j.vetmic.2010.08.024.
28. Nogami K, Maeda M. Bacteria As Biocontrol Agents for Rearing Larvae of the Crab Portunus-Trituberculatus. Can J Fish Aquat Sci. 1992;49(11):2373-2376. doi:10.1139/f92-261.
29. Rengpipat S., Rukpratanporn S, S. Piyatiratitivorakul and PM. Probiotics in Aquaculture: a case study of probiotics for larvae of the black tiger shrimp (Penaeus monodon). In: Flegel TW, ed. Advances in Shrimp Biotechnology. National Center for Genetic Engineering and Biotechnology, Bangkok (TH); 1998:52-56.
30. Jha AK. Probiotics: An effective food supplemnt in shrimp culture. J Bangladesh Acad Sci. 2014;38:127-130. http://www.banglajol.info/ index.php/JBAS/article/viewFile/21336/14649.
31. Lara-Flores M, Olvera-Novoa MA, Guzmán-Méndez BE, López-Madrid W. Use of the bacteria Streptococcus faecium and Lactobacillus acidophilus, and the yeast Saccharomyces cerevisiae as growth promoters in Nile tilapia (Oreochromis niloticus). Aquaculture. 2003;216(1-4):193-201. doi:10.1016/S0044-8486(02)00277-6.
32. Irianto A, Austin B. Use of dead probiotic cells to control furunculosis in rainbow trout, Onchorhynchus mykiss. J Fish Dis. 2003;26:59-62.
33. Lazado CC, Caipang CMA, Kiron V. Enzymes from the gut bacteria of Atlantic cod, Gadus morhua and their influence on intestinal enzyme activity. Aquac Nutr. 2012;18(4):423-431. doi:10.1111/j.1365-2095.2011.00928.x.
34. Johansson MW, Keyser P, Sritunyalucksana K, Söderhäll K. Crustacean hemocytes and haematopoiesis. Aquaculture. 2000;191:45-52.
35. Leonel Ochoa-Solano J, Olmos-Soto J. The functional property of Bacillus for shrimp feeds. Food Microbiol. 2006;23(6):519-525. doi:10.1016/j.fm.2005.10.004.
36. Chiu C-H, Guu Y-K, Liu C-H, Pan T-M, Cheng W. Immune responses and gene expression in white shrimp, Litopenaeus vannamei, induced by Lactobacillus plantarum. Fish Shellfish Immunol. 2007;23(2):364-377. doi:10.1016/j.fsi.2006.11.010.
37. Vieira FN, Buglione CC, Mouriño JPL, et al. Effect of probiotic supplemented diet on marine shrimp survival after challenge with Vibrio harveyi. Arq Bras Med Veterinária e Zootec. 2010;62(3):631-638. doi:10.1590/S0102-09352010000300019.
38. Brousseau JP, Talbot G, Beaudoin F, Lauzon K, Roy D, Lessard M. Effects of probiotics Pediococcus acidilactici strain MA18/5M and Saccharomyces cerevisiae subsp. boulardii strain SB-CNCM I-1079 on fecal and intestinal microbiota of nursing and weanling piglets. J Anim Sci. 2015;93(11):5313-5326. doi:10.2527/jas.2015-9190.
39. Castex M, Lemaire P, Wabete N, Chim L. Effect of probiotic Pediococcus acidilactici on antioxidant defences and oxidative stress of Litopenaeus stylirostris under Vibrio nigripulchritudo challenge. Fish Shellfish Immunol. 2010;28(4):622-631. doi:10.1016/j.fsi.2009.12.024.
40. Gullian M, Thompson F, Rodriguez J. Selection of probiotic bacteria and study of their immunostimulatory effect in Penaeus vannamei. Aquaculture. 2004;233(1-4):1-14. doi:10.1016/j.aquaculture.2003.09.013.
41. Lakshmi B, Viswanath B, Sai Gopal DVR. Probiotics as antiviral agents in shrimp aquaculture. J Pathog. 2013;2013:424123. doi:10.1155/2013/424123.
42. Thompson J, Gregory S, Plummer S, Shields RJ, Rowley AF. An in vitro and in vivo assessment of the potential of Vibrio spp. as probiotics for the Pacific White shrimp, Litopenaeus vannamei. J Appl Microbiol. 2010;109(4):1177-1187. doi:10.1111/j.1365-2672.2010.04743.x.
43. Bricknell I, Dalmo RA. The use of immunostimulants in fish larval aquaculture. Fish Shellfish Immunol. 2005;19(5 SPEC. ISS.):457-472. doi:10.1016/j.fsi.2005.03.008.
44. Meena DK, Das P, Kumar S, et al. Beta-glucan: An ideal immunostimulant in aquaculture (a review). Fish Physiol Biochem. 2013;39(3):431-457. doi:10.1007/s10695-012-9710-5.
45. Raa J. Immune modulation by non-digestible and non-absorbable beta-1,3/1,6-glucan. Microb Ecol Health Dis. 2015;26:1-4.
46. Dawood MAO, Koshio S, Ishikawa M, et al. Dietary supplementation of β-glucan improves growth performance, the innate immune response and stress resistance of red sea bream, Pagrus major. Aquac Nutr. 2016. doi:10.1111/anu.12376.
47. Kumar V, Roy S, Meena DK, Sarkar UK. Application of Probiotics in Shrimp Aquaculture: Importance, Mechanisms of Action, and Methods of Administration. Rev Fish Sci Aquacululture. 2016;24:342-368.
48. Pais-Chanfrau JM, Trujillo-Toledo LE. Optimization of culture medium for large-scale production of heterologous proteins in Pichia pastoris to be used in nanoscience and other biotechnological fields. Biol Med. 2016;8(3). doi:10.4172/0974-8369.1000279.
49. Mahious AS, Gatesoupe FJ, Hervi M, Metailler R, Ollevier F. Effect of dietary inulin and oligosaccharides as prebiotics for weaning turbot, Psetta maxima (Linnaeus, C. 1758). Aquac Int. 2006;14(3):219-229. doi:10.1007/s10499-005-9003-4.
50. Fooks LJ, Fuller R, Gibson GR. Prebiotics, probiotics and human gut microbiology. Int Dairy J. 1999;9(1):53-61. doi:10.1016/S0958-6946(99)00044-8.
51. Sang HM., Ky LT, Fotedar R. Dietary supplementation of mannan oligosaccharide improves the immune responses and survival of marron, Cherax tenuimanus when challenged with different stressors. Fish Shellfish Immunol. 2009;27:341-348.
52. Sang HM, Fotedar R. Effects of dietary β-1,3-glucan on the growth, survival, physiological and immune response of marron, Cherax tenuimanus. Fish Shellfish Immunol. 2010;28:957-960.
53. Bailey JS, Blankenship LC, Cox NA. Effect of fructooligosaccharide on Salmonella colonization of the chicken intestine. Poult Sci. 1991;70(12):2433-2438. doi:10.3382/ps.0702433.
54. Carbone D, Faggio C. Importance of prebiotics in aquaculture as immunostimulants. Effects on immune system of Sparus aurata and Dicentrarchus labrax. Fish Shellfish Immunol. 2016;54:172-178. doi:10.1016/j.fsi.2016.04.011.
55. Song SK, Beck BR, Kim D, et al. Prebiotics as immunostimulants in aquaculture: A review. Fish Shellfish Immunol. 2014;40(1):40-48. doi:10.1016/j.fsi.2014.06.016.
56. Mussatto SI, Mancilha IM. Non-digestible oligosaccharides: A review. Carbohydr Polym. 2007;68(3):587-597. doi:10.1016/j.carbpol.2006.12.011.
57. Li P, Burr GS, Gatlin DM, et al. Dietary supplementation of short-chain fructooligosaccharides influences gastrointestinal microbiota composition and immunity characteristics of Pacific white shrimp, Litopenaeus vannamei, cultured in a recirculating system. J Nutr. 2007;137(12):2763-2768. doi:137/12/2763 [pii].
58. Gatlin III DM, Li P, Wang X, Burr GS, Castille F, Lawrence AL. Potential application of prebiotics in aquaculture. VIII Simp Int Nutr Acuicola. 2006:371-376.
59. Gibson GR. Dietary modulation of the human gut microflora using the prebiotics oligofructose and inulin. J Nutr. 1999;129(7 Suppl):1438S-41S.
60. Gibson GR. Nutritional and Health Benefits of Inulin and Oligofructose Dietary Modulation of the Human Gut Microflora Using the Prebiotics Oligofructose and Inulin 1. Pathology. 1999;129(7 Suppl):1438-1441. http://www.ncbi.nlm.nih.gov/pubmed/10395616.
61. Trujillo L, Granda S, Ordoñez M, Ramos T, Avalos R. Detection of short chain inulin (FOS)-fermenting microorganisms useful in food and health science. African J Biotecnol. 2015;14(38):2759-2764. doi:10.5897/AJB2015.14852 I.
62. Grisdale-Helland B, Helland SJ, Gatlin DM. The effects of dietary supplementation with mannanoligosaccharide, fructooligosaccharide or galactooligosaccharide on the growth and feed utilization of Atlantic salmon (Salmo salar). Aquaculture. 2008;283(1-4):163-167. doi:10.1016/j.aquaculture.2008.07.012.
63. Li P, Gatlin DM. Dietary brewers yeast and prebiotic Grobiotic® AE influencegrowth performance immnue responses and resistance of hybrid striped sea bass (Moronechrysops x M. saxatilis) to streptococcus iniae infection. Aquaculture. 2004;231:445-456.
64. Refstie S, Bakke-McKellep AM, Penn MH, Sundby A, Shearer KD, Krogdahl Å. Capacity for digestive hydrolysis and amino acid absorption in Atlantic salmon (Salmo salar) fed diets with soybean meal or inulin with or without addition of antibiotics. Aquaculture. 2006;261(1):392-406. doi:10.1016/j.aquaculture.2006.08.005.
65. Mahious AS, Ollevier F. Probiotics and Prebiotics in Aquaculture. In: 1st Regional Workshop on Techniques for Enrichment of Live Food for Use in Larviculture-2005, AAARC, Urmia, Iran. ; 2005:67.
66. Yilmaz, E., Genc MA, Genc E. Effects of dietary mannanoligosaccharides on growth, body composition, and intestine and liverhistology of rainbow trout, Oncorhynchus mykiss. Isr J Aquac. 2007;59:182-188.
67. Genc MA, Aktas M, Genc E, Yilmaz E. Effects of dietary mannan oligosaccharide on growth, body composition and hepatopancreas histology of Penaeus semisulcatus (de Haan 1844). Aquac Nutr. 2007;13(2):156-161. doi:10.1111/j.1365-2095.2007.00469.x.
68. Akrami A, Hajimoradloo A, Matinfar A, Abedian Kinai A. Effect of dietary prebiotic Inulin on growth performance, intestinal microflora, body composition and hematological performance of juvenile Beluga Huso huso (Linnaeus, 1758). J World Aquac Soc. 2010;40:771-779.
69. Sequeira T, Tavares D, Arala-Chaves M. Evidence for circulating hemocyte proliferation in the shrimp Penaeus japonicus. Dev Comp Immunol. 1996;20(2):97-104. doi:10.1016/0145-305X(96)00001-8.
70. Perazzolo LM, Barracco MA. The prophenoloxidase activating system of the shrimp Penaeus paulensis and associated factors. Dev Comp Immunol. 1997;21(5):385-395. doi:10.1016/S0145-305X(97)00022-0.
71. Fukasaka M, Asari D, Kiyotoh E, et al. A lipopolysaccharide from pantoea agglomerans is a promising adjuvant for sublingual vaccines to induce systemic and mucosal immune responses in Mice via TLR4 pathway. PLoS One. 2015;10(5). doi:10.1371/journal.pone.0126849.
72. van de Braak CBT, Botterblom MH a, Liu W, Taverne N, van der Knaap WPW, Rombout JHWM. The role of the haematopoietic tissue in haemocyte production and maturation in the black tiger shrimp (Penaeus monodon). Fish Shellfish Immunol. 2002;12(3):253-272. doi:10.1006/fsim.2001.0369.
73. Okumura T. Effects of lipopolysaccharide on gene expression of antimicrobial peptides (penaeidins and crustin), serine proteinase and prophenoloxidase in haemocytes of the Pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2007;22(1):68-76. doi:10.1016/j.fsi.2006.03.013.
74. Ng TH, Chang SH, Wu MH, Wang HC. Shrimp hemocytes release extracellular traps that kill bacteria. Dev Comp Immunol. 2013;41(4):644-651. doi:10.1016/j.dci.2013.06.014.
75. Rungrassamee W, Maibunkaew S, Karoonuthaisiri N, Jiravanichpaisal P. Application of bacterial lipopolysaccharide to improve survival of the black tiger shrimp after Vibrio harveyi exposure. Dev Comp Immunol. 2013;41(2):257-262. doi:10.1016/j.dci.2013.05.021.
76. Zhang XW, Wang Y, Wang XW, Wang L, Mu Y, Wang JX. A C-type lectin with an immunoglobulin-like domain promotes phagocytosis of hemocytes in crayfish Procambarus clarkii. Sci Rep. 2016;6:29924.
77. Zhang S, Shi L, Yang Q-H, et al. Molecular characterization and functional analysis of Cactin gene from Litopenaeus vannamei. Fish Shellfish Immunol. 2014;41(2):608-617. doi:10.1016/j.fsi.2014.10.014.
78. Chen YY, Chen JC, Lin YC, et al. Endogenous molecules induced by a Pathogen-Associated Molecular Pattern (PAMP) elicit innate immunity in shrimp. PLoS One. 2014;9(12). doi:10.1371/journal.pone.0115232.
79. Xian JA, Zhang XX, Guo H, Wang DM, Wang AL. Cellular responses of the tiger shrimp Penaeus monodon haemocytes after lipopolysaccharide injection. Fish Shellfish Immunol. 2016;54:385-390.
80. Akhter N, Wu B, Memon AM, Mohsin M. Probiotics and prebiotics associated with aquaculture: A review. Fish Shellfish Immunol. 2015;45(2):733-741. doi:10.1016/j.fsi.2015.05.038.
81. Huynha T-G, Shiua Y-L, Nguyenb, Thanh-Phuong Truongb Q-P, Chenc J-C, Chun-Hung L. Current applications, selection, and possible mechanisms of actions of synbiotics in improving the growth and health status in aquaculture: A review. Fish Shellfish Immunol. 2017;In Press,.
82. Widanarni, Tanbiyaskur. Application of probiotic, prebiotic and synbiotic for the control of streptococcosis in tilapia Oreochromis niloticus. Pakistan J Biol Sci. 2015;18(2):59-66. doi:10.3923/pjbs.2015.59.66.
83. Lazado CC, Lacsamana JI, Caipang CMA. Mechanisms of probiotic actions in shrimp: Implications to tropical aquaculture. In: Caipang CMA, Bacano-Maningas MBI, Fagutao FF, eds. Biotechnological Advances in Shrimp Health Management in the Philippines. Kerala, India: Research Signpost; 2015.
Recibido: febrero 2017
Aprobado: Mayo 2017
Luis E. Trujillo1*, Leonor Rivera2,3, Eugenio Hardy1,4, Erika M. Llumiquinga1, Francisco Garrido1, Jesús A. Chávez3, Victor H. Abril1, José M. País-Chanfrau5
1Universidad de las Fuerzas Armadas ESPE. Life science department. Industrial Biotechnology and Bioproducts Research Group, Center for Nanotechnology and Nanosciences CENCINAT, Quito, Pichincha, Ecuador
2Universidad Técnica de Machala, Machala, El Oro, Ecuador
3Unidad de Posgrado de la Universidad Nacional Mayor de San Marcos-Perú
4Biotechnology Laboratory, Study Center for Research and Biological Evaluations, Institute of Pharmacy and Foods, Havana University, Cuba
5Universidad Tecnica del Norte UTN. FICAYA. Ave. 17 julio 5-21, Ibarra 119, Imbabura, Ecuador
*Correspondence author: [email protected] (L.E. Trujillo)