2022.07.02.44
Files > Volume 7 > Vol 7 No 2 2022

Saad Sabah Fakhry1, Zainab Noori Hammed2, Wasan Abdul - elah Bakir 3, Bahaa Abdullah Laftaah ALRubaii4*
1 Ministry of Higher education and scientific research, science and technology-Environment and water Directorate food contamination research center, Baghdad, Iraq.
2AlHikma University College, Baghdad, IRAQ
3 Microbiology department/ Bacteriology College of medicine / Mustansiriyah University
4 University of Baghdad, Baghdad, College of Science, Department of Biology, Baghdad, Iraq
*Corresponding author email is: [email protected]
Available from: http://dx.doi.org/10.21931/RB/2022.07.02.44
ABSTRACT
Staphylococcus aureus causes mastitis in dairy cows, lambs, goats, and skin disorders in pigs and other animals. S. aureus causes localized purulent infections that affect soft tissues, bones, and other organs in humans. Using restriction patterns, the researchers want to isolate and identify methicillin-resistant Staphylococcus aureus (MRSA) strains from cattle and humans. They also hope to assess their genetic relatedness by comparing the mecA1 and mecA2 gene sequence discrepancies. Animals (223 strains) and people have been used to acquire S. aureus strains for study (83). The E-test was used to assess whether or not the bacteria were resistant to methicillin. The mecA1 and mecA2 genes were identified by using pulsed-field gel electrophoresis (PFGE) to analyze DNA restriction patterns. The results were shown. S. aureus strains from animals and men were resistant to methicillin in 32 (14.34 %) and 53 (63.8 %), respectively. PFGE was used to determine the differences between human and veterinary pathology strains. Two strains of bacteria collected from animals were discovered to be identical; nevertheless, microorganisms recovered from humans were found to be significantly similar to the bacteria recovered from animals. Both human and veterinary pathology were implicated in the development of methicillin resistance. The MRSA strains found in humans were much more significant than those found in animals. The strains recovered from animals exhibited a high degree of genetic heterogeneity. Still, the enormous number of indistinguishable bacteria in humans leads one to believe that a dominant clone is present. When it comes to the molecular characterization of MRSA isolates, PFGE might be regarded as the gold standard.
Keywords. Animals, Human, MRSA, PFGE, Staphylococcus, mecA genes
INTRODUCTION
A report on the occurrence of methicillin-resistant S. aureus (MRSA) in medicine was published soon after the antibiotic methicillin was introduced into clinical practice to treat infections caused by these germs 1. Characteristics of staphylococci that increase their pathogenicity include their propensity to colonize and multiply actively. When the infection takes hold, contributing risk factors include the slow healing of certain recurring infections, the use of lengthy antibiotic treatments, the use of catheters, and, once again, the presence of open wounds2. In dairy cows, S.aureus is one of the most common agents responsible for mastitis 3, 4, sheep and goats and is often involved in skin infections in pigs and other animal species 5. S. aureus is the leading etiological agent of localized purulent infections affecting soft tissues, bones, and other organs in humans. It is mainly responsible for infections of natural exogenous according to its ability to spread in the environment, thus causing epidemics in closed communities such as hospitals, homes in care and schools 6,7,8. Under some situations, the microbe demonstrates a high virulence capability, capable of exacerbating the inflammatory process till the host dies. Methicillin-resistant strains have been recorded in dogs 9, horses, and dairy cows after treating staphylococcal infections with methicillin and -lactams in general 4. In humans, the MRSA strains are fearsome pathogens, especially in the hospital field, and countless epidemics are reported in the bibliography 7 . It has recently been proven that the transmission routes can be mainly two, one direct from patient to patient and the other, indirect, through healthcare professionals 10, 7. Methicillin resistance arises due to the presence of the mecA genes those code for a Penicillin Binding Protein (PBP2a), which has a low affinity for β-lactam compounds 11. Veterinary pathology has long recognized that resistance to methicillin and oxacillin, a closely related antibiotic extensively used in veterinary pathology, is often linked with analogous behavior against a broad spectrum of antibiotics 12 to compromise therapeutic interventions, which, concerning bovine mastitis, represent the most effective method for controlling the disease. Now the 1960s, MRSA isolates in people have been recorded in European hospitals, and the disease has since expanded to non-hospital settings as well 13 so much so that, At the moment, MRSA jambs account for 25 percent of the strains of S. aureus that cause nosocomial infections in hospitals. In this work, the goal was to determine the existence of isolated MRSA strains in both human and animal samples and submit these jambs for molecular examination of chromosomal DNA through PFGE.
MATERIALS AND METHODS
Bacterial isolates
From the mammary secretion of cows with mastitis subclinical, 223 S. aureus isolates have been obtained. Of patients admitted to different wards of the hospital, 83 were selected from the isolates of S. aureus. The pathological material was sown in Nutrient Agar with the addition of 5% sheep blood and in Mannitol Salt Agar. The isolated microorganisms were identified by coagulase and Dnase tests, API Staph.
Methicillin-resistance
Mueller Hinton Agar was used to cultivate the isolates.Following that, 4-5 isolated colonies were collected and planted in Nutrient Broth up to a turbidity of 0.5 McFarland in 2-8 hours at 37°C till the turbidity of 0.5 McFarland was reached. For the determination of the Minimum Inhibiting Concentration (MIC) expressed in micrograms per milliliter (g/ml), an aliquot of 0.01 ml was diluted in 10 ml of saline solution, and 3 ml were distributed on the surface of a plate Mueller Hinton Agar 14 on which they have placed the nitrocellulose strips containing Methicillin E-test for the determination of the Minimum Inhibiting Concentration. Following incubation at 35°C for 24 hours, S. aureus jambs were tested for resistance (MRSA). Those that showed an equal or more significant inhibition at µg / ml were designated as resistant (MRSA). As a result of PCR15, the existence of the mecA gene has been identified. The QIAmp tissue Kit was used to perform the DNA purification.The primers mecA1 (5'-AAA ATC GAT GGT AAA GGTTGG C) and mecA2 (5'- AGT TCT GCA GTA CCGGAT TTG C) highlight a product of PCR of 533bp 16. Ungeheuer's method of DNA amplification was used 17. DNA electrophoresis was carried out on a gel containing 2 % agarose, after which it was colored with ethidium bromide and photographed under the influence of ultraviolet light.
Pulsed-field gel electrophoresis (PFGE)
The macro-restriction approach with Sma I described the MRSA jambs, which were then resolved by pulsed electrophoresis18. In 10 ml of Trypticase Soy Broth at 37°C for 24 hours, the bacteria were grown before being centrifuged at 2000 g for 10 minutes, and the pellet was resuspended in PIV buffer. To manufacture the plugs, an aliquot of 0.5 ml of the latter suspension was combined with an equal quantity of Incert Agarose (low melting) 1 % to form a homogeneous mixture. Each plug received one milliliter of lysis solution, and they were incubated at 37°C for 24 hours in agitation. The lysis solution was replaced with a protein digestion buffer and then incubated at 50°C for 48 hours under agitation. The plugs were washed with TE and PMSF and three more times with TE. Later iplugs were subjected to enzymatic digestion in a mixture of H2O sterile, spermidine (1 mM), SmaI 100U. After boiling for around 24 hours at 25°C in a final volume of 250 mL, the solution was utilized for pulsed electrophoresis on a 1 percent Fast Lane agarose gel at 14°C in TBE buffer auto-algorithm for 24 hours. Lambda Ladder was the standard that was utilized. The PFGE was done out using the CHEF Mapper system, which may be found here. To build a similarity dendrogram18, the patterns electrophoretic were captured using a camera in the Molecular program Analyst Finger Printing and analyzed using the MA Finger Printing. The similarity coefficients (SAB) were estimated using software that was not modified in any way. From 0 to 1, the SAB indicated whether or not the patterns were connected, with 0 indicating that they were not related and one suggesting that they were tightly associated.
RESULTS
The chemical and metabolic properties of the 306 S. aureus isolates were consistent throughout the group. Of the 223 jambs studied, 32 (14.34 percent) were found to be resistant to methicillin, but of the 83 jambs investigated that were of human origin, 53 (63.8 percent) were found to be MRSA. Forty MRSA and twenty MSSA jambs, derived in equal parts from human and veterinary illness, in which the presence of the mecA gene has been validated by PCR to verify the findings obtained with the E-test and Sceptor System, have been subjected to PFGE. As a consequence of the PFGE data, the investigation of the genetic similarity of the MRSA jambs, which was shown using a dendrogram (Figs. 1 and 2), revealed that the strains of human and veterinary origin were distinct. The blocks that arrived showed a significant degree of resemblance among themselves; however, just two strains of animal origin showed identical results compared to the unions that came (Table 1).
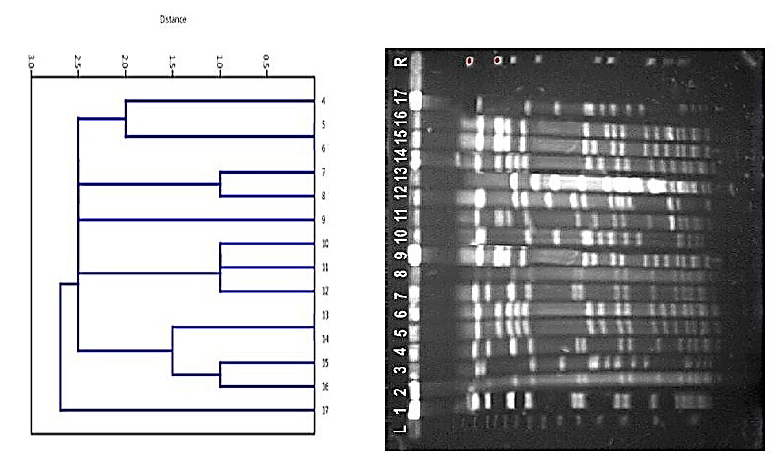
Figure 1. Staphylococcus aureus (MRSA) of human origin: similarity dendrogram.
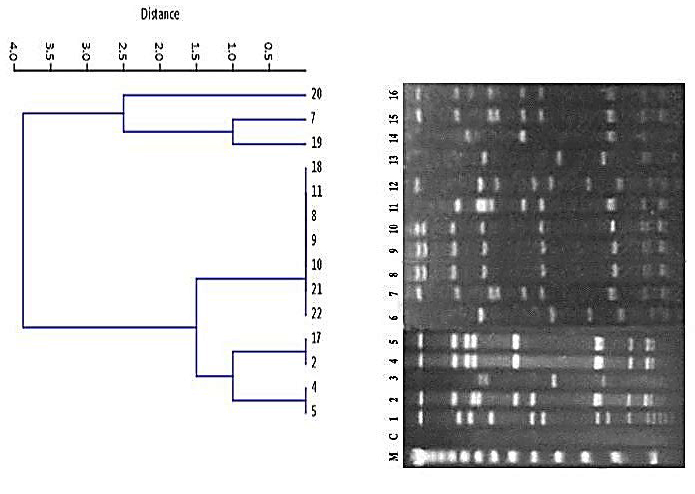
Figure 2. Staphylococcus aureus (MRSA) of animal origin: similarity dendrogram.
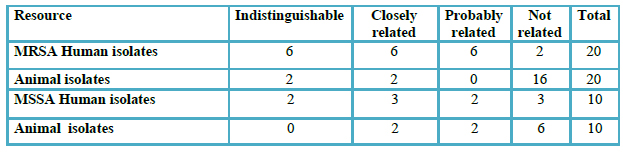
Table 1: Staphylococcus aureus: genomic similarity of the isolates from human and animal pathology.
DISCUSSION
Staphylococcus aureus is one of the most prevalent germs found in nature and one of the most responsible for illness, both in people and in animals. It can produce infections that are difficult to treat with conventional therapy19. To adapt to the increased use of antibiotics, S. aureus has modified its genomic structure and made the PBP2a protein via the mecA genes, which has a low affinity for -lactam antibiotics, leading in the appearance of methicillin-resistant jambs 20, 18. One of the issues that must be addressed is selecting a technique for identifying staphylococcal jambs. The identification of MRSA strains is difficult, even with the international set of phages, and it was therefore deemed appropriate to replace conventional methods with molecular biology techniques, such as PFGE or polymerase chain reaction, which, when performed with the primers mecA1 and mecA2, results in the identification of a sequence of 533 base pairs, specific for MRSA jambs14. Since it has been demonstrated in human pathology that MRSA jambs from different continents are classified as indistinguishable, it was deemed appropriate to compare jambs of human and veterinary origin, in the same territorial context, to verify if they had been realized under the same conditions. Although the jambs of animal origin originated from farms in different geographies, this does not excuse the lack of genetic similarity between them, as MRSA jambs considered unrecognizable have a genetic similarity of 80% or greater 21,22. When a comparison analysis was conducted between the 40 MRSA jambs and the 20 MSSA jambs, it was discovered that there is no genetic similarity between them, just as there were no commonalities between the MRSA jambs from veterinary and human pathology. However, only two MRSA jambs of veterinary origin could be identified as genetically identical and two other closely related ones.
In contrast, in human pathology, the genetically similar jambs (n.6) and closely related jambs (n. 6) suggested the presence of a dominant strain in the degree to which there is a close correlation between strains isolated over time. Aside from the genetic similarities, the difficulties associated with methicillin resistance impact veterinary and human illnesses similarly. Pulsed electrophoresis is widely recognized as the gold standard for molecular typing of a broad variety of microorganisms. It has been shown to be highly selective and equivalent, if not better, to other approaches for a large number of harmful bacteria16,18,4. The findings of the investigation, despite being constrained by a small number of jambs, confirm the presence of MRSA in cattle and demonstrate the high level of genomic variability found within methicillin-resistant strains, which suggests that these bacteria could spread from animals to the environment through contact with other animals other than humans 20 . As an additional indication of the complex somatic structure of S. aureus, there is an inverse relationship between the homogeneity shown by chemical-metabolic analysis and genomic inequality, which is the element that best defines the bacterium.
CONCLUSION
MRSA isolates from several hospital wards have been studied. The results indicate a significant degree of similarity between certain jambs, which suggests the presence of a typical clone from which they might originate the isolates. This clone has been present at the hospital for at least 15 years. It has caused periodic epidemics (data not published), leading some to speculate that it has colonized the healthcare workers, who would then act from tanks and vehicles due to the outbreak. The genetic analysis of MRSA strains highlights the necessity for adequate procedures to be taken to monitor clinical isolates so that they may act effectively to stop the epidemiological cycle, as shown by the research.
Acknowledgment: Thanks going for all who support us.
Conflict between authors: No conflict
Funds: self by authors
REFERENCES
1. Barber M. Methicillin-resistant staphylococci. Journal of clinical pathology. 1961; 14(4):385
2. Meakins JL, Wicklund B, Forse RA, McLean AP. The surgical intensive care unit: current concepts in infection. Surgical Clinics of North America. 1980 1;60(1):117-32.
3. Devriese LA, Hommez J. Epidemiology of methicillin-resistant Staphylococcus aureus in dairy herds. Research in veterinary science. 1975; 19(1):23-27.
4. Zadoks R, Van Leeuwen W, Barkema H, Sampimon O, Verbrugh H, Schukken YH, Van Belkum A. Application of pulsed-field gel electrophoresis and binary typing as tools in veterinary clinical microbiology and molecular epidemiologic analysis of bovine and human Staphylococcus aureus isolates. Journal of clinical microbiology. 2000; 38(5):1931-1939.
5. Scott GM, Thomson R, Malone-Lee J, Ridgway GL. Cross-infection between animals and man: possible feline transmission of Staphylococcus aureus infection in humans?. Journal of Hospital Infection. 1988; 12(1):29-34.
6. Allen KD, Anson JJ, Parsons LA, Frost NG. Staff carriage of methicillin-resistant Staphylococcus aureus (EMRSA 15) and the home environment: a case report. Journal of Hospital Infection. 1997; 35(4):307-311.
7. Emmerson M. Nosocomial staphylococcal outbreaks. Scandinavian journal of infectious diseases. Supplementum. 1994; 93:47-54.
8. Fraise AP, Mitchell K, O’BRIEN SJ, Oldfield K, Wise R. Methicillin-resistant Staphylococcus aureus (MRSA) in nursing homes in a major UK city: an anonymized point prevalence survey. Epidemiology & Infection. 1997; 118(1):1-5.
9. Tomlin J, Pead MJ, Lloyd DH, Howell S, Hartmann F, Jackson HA, Muir PJ. Methicillin‐resistant Staphylococcus aureus infections in 11 dogs. Veterinary Record. 1999; 144(3):60-64.
10. Cox RA, Conquest C. Strategies for the management of healthcare staff colonized with epidemic methicillin-resistant Staphylococcus aureus. Journal of Hospital Infection. 1997; 35(2):117-127.
11. Hussain Z, Stoakes L, Garrow S, Longo S, Fitzgerald V, Lannigan R. Rapid detection of mecA-positive and mecA-negative coagulase-negative staphylococci by an anti-penicillin binding protein 2a slide latex agglutination test. Journal of Clinical Microbiology. 2000;38(6):2051-2054.
12. Barie PS. Antibiotic-resistant gram-positive cocci: implications for surgical practice. World journal of surgery. 1998; 22(2):118-126.
13. Moreno F, Crisp C, Jorgensen JH, Patterson JE. Methicillin-resistant Staphylococcus aureus as a community organism. Clinical Infectious Diseases. 1995; 21(5):1308-1312.
14. Kampf G, Weist K, Swidsinski S, Kegel M, Rüden H. Comparison of screening methods to identify methicillin-resistantStaphylococcus aureus. European Journal of Clinical Microbiology and Infectious Diseases. 1997;16(4):301-307.
15. Murakami K, Minamide W, Wada K, Nakamura E, Teraoka H, Watanabe S. Identification of methicillin-resistant strains of staphylococci by polymerase chain reaction. Journal of clinical microbiology. 1991; 29(10):2240-2244.
16. Murray PR, Baron EJ, Pfaller MA, Tenover FC, Yolken RH, Morgan DR. Manual of Clinical Microbiology (6th edn). Trends in microbiology. 1995; 3(11):449-449.
17. Ungeheuer J, Wilms S, Sietzen W. mecA-Gen bei Staphylokokken—vereinfachter Nachweis mit der PCR. Chemother J. 1994; 1:31-35.
18. Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH, Swaminathan B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. Journal of clinical microbiology. 1995; 33(9):2233-2239.
19. Cefai C, Ashurst S, Owens C. Human carriage of methicillin-resistant Staphylococcus aureus linked with pet dog. The Lancet. 1994 ;344(8921):539-540.
20. Shukra AM, Sridevi NV, Chandran D, Maithal K. Production of recombinant antibodies using bacteriophages. European Journal of Microbiology and Immunology. 2014; 4(2):91-98.
21. Mato R, Sanches IS, Venditti M, Platt DJ, Brown A, Chung M, De Lencastre H. Spread of the multiresistant Iberian clone of methicillin-resistant Staphylococcus aureus (MRSA) to Italy and Scotland. Microbial Drug Resistance. 1998;4(2):107-112.
22. Teixeira LA, Resende CA, Ormonde LR, Rosenbaum R, Figueiredo AM, De Lencastre H, Tomasz A. Geographic spread of epidemic multiresistant Staphylococcus aureus clone in Brazil. Journal of clinical microbiology. 1995; 33(9):2400-2404.
Received: 14 January 2022 / Accepted: 29 March 2022 / Published:15 May 2022
Citation. Sabah Fakhry S, Noori Hammed Z , Abdul - elah Bakir W, Abdullah Laftaah ALRubaii B . Identification of methicillin-resistant strains of Staphylococcus aureus isolated from humans and food sources by Use of mecA 1 and mecA 2 genes in Pulsed-field gel electrophoresis (PFGE (technique. Revis Bionatura 2022;7(2) 44. http://dx.doi.org/10.21931/RB/2022.07.02.44