2021.06.03.3
Files > Volume 6 > Vol 6 No 3 2021
INVESTIGATION / RESEARCH
Safety and immunogenicity in piglets of two immunization schedules initiated at two or three weeks of age with PorvacÒ, a classical swine fever subunit marker vaccine.

Aymé Oliva-Cárdenas1, Fé Fernández-Zamora 2, Elaine Santana-Rodríguez1, Yusmel Sordo-Puga1, Milagros de la C. Vargas-Hernández1, María P. Rodríguez-Moltó1, Danny Pérez-Pérez1, Talia Sardina-González1,Carlos A. Duarte1, Avelina León-Goñi 2, Diurys Blanco -Gámez2, Francisco Contreras-Pérez2, Odalys Valdés-Faure2, Rosmery Hernández-Prado1, Eric Acosta-Lago2, Ileana Sosa-Testé2, Marisela F. Suárez-Pedroso1
Available from: http://dx.doi.org/10.21931/RB/2021.06.03.3
ABSTRACT
Classical swine fever is a highly contagious viral disease with a significant impact on food production worldwide. It currently represents one of the main limitations for the development of the pig industry in Cuba. PorvacÒ is a subunit marker vaccine that confers a very rapid onset of protection. Since there are different production systems in pig breeding, readjustments in the vaccination program are often required. This study compares the safety and efficacy in piglets of two vaccination schedules with PorvacÒ (0-2 weeks and 0-3 weeks), initiated at two or three weeks of age. Clinical monitoring was conducted, and a neutralization peroxidase-linked assay was used to measure the neutralization titers. All immunization regimens were safe and well-tolerated, without local or systemic adverse reactions in the vaccinated animals. Geometric mean neutralizing antibody titers higher than 1/1500 were detected in all groups during the six months of the trial. One month after the second immunization, piglets primed at two weeks of age, and boostered three weeks later, developed significantly higher neutralization titers (1/15644) compared to those vaccinated at a similar age but with a two-week interval between doses (1/5760). However, no significant differences in the titers were found three and six months after vaccination among the four regimens. In summary, all the variants studied are effective, but it is recommended to start vaccination at two weeks old, with the second dose at either two or three weeks later, depending on the production system and the purpose of the farm.
Keywords. Classical swine fever; E2CD154; subunit vaccine; piglets; neutralizing antibodies
INTRODUCTION
Classical swine fever (CSF) is an infectious disease with the most significant economic impact on the swine industry in Cuba and many other countries. The disease is caused by the classical swine fever virus (CSFV), an enveloped, single-stranded RNA Pestivirus. Due to its devastating effects, CSF has been listed as a notifiable disease by the World Organization for Animal Health (OIE)1. In endemic areas, prophylactic vaccination is commonly used to limit the effects of the disease or as a first step within a general program to control and eradicate the virus. The modified live vaccines (MLV) are the most widely used in CSF endemic regions and have effectively controlled the disease in several countries such as Brazil, Argentina, and Uruguay.
On the other hand, Cuba is one CSF endemic country where the virus has been emerging for more than 20 years despite the use of an attenuated C strain MLV 2-4. The lack of discrimination between infected and vaccinated animals, the requirement for a cold chain of distribution, and the interference of maternal-derived neutralizing antibodies (MDNA) are the main drawbacks of MLV, which have limited its use in disease-free regions. Several subunit vaccines have been developed to overcome these drawbacks, but they exhibit a late onset of protection as compared to MLV and provide insufficient protection against vertical transmission5-8.
PorvacÒ is a Cuban subunit marker vaccine against CSFV, which is based on a recombinant chimeric antigen comprising the E2 protein of CSFV and the molecular adjuvant CD154. This vaccine has been safe and capable of inducing an unusually rapid onset of protection against a challenge with a highly pathogenic CSFV strain, which is similar to the one described for MLV 9-11. PorvacÒ induces both high neutralizing antibodies (NAb) titers and cell-mediated immune response and can interfere with the transmission of the virus from pregnant sows to their offspring 12.
PorvacÒ has been registered in Cuba and is been currently used in both, large state-owned pig production units and small private farms. Since there are different production systems in swine breeding, readjustments in the immunization schedules must make them compatible with those systems. The present study aimed to compare the safety and efficacy of two different immunization schedules with PorvacÒ (0-2 weeks and 0-3 weeks), starting at either two or three weeks after birth. These experiments must provide the scientific basis for a more flexible approach in terms of vaccination schedules for the different production systems.
METHODS
PorvacÒ vaccine
The vaccine preparation was obtained from the stable HEK 293 cell line (ATCC CRL1573), which expressed the CSFV E2 antigen fused to the porcine CD154 protein. It was produced under Good Manufacturing Practices in a certified production area of the Center for Genetic Engineering of Camaguey, Cuba, and formulated as a water-in-oil formulation with Montanide ISA50V2 (SEPPIC, France) at a final concentration of 25 μg of E2-CD154 antigen/ mL.
Experimental animals
The study included the litters from four unvaccinated CSFV-negative sows (Duroc x Yorkshire crossbred swine). The piglets were kept with their mothers until weaning, between 33 and 42 days, each group in a separate experimental room. Trials were carried out under appropriate high containment conditions following the animal welfare regulations and standards according to Good Clinical Practices 13-15. The study was approved and supervised by the Committee for Care and Use of Laboratory Animals (CICUAL) of the Center for the Production of Laboratory Animals (CENPALAB).
The general clinical conditions of the animals were evaluated during the immunization period, especially the occurrence of adverse reactions at the injection site. The rectal temperature was measured immediately before the first vaccination and 72 hours after each vaccine administration. Blood samples were taken from the ophthalmic venous sinus in the absence of anticoagulant, previous to the first immunization (T0) and at one, three, and six months after the second immunization (T1, T3, and T6, respectively). Hematological and biochemical analyses were conducted at the analytical laboratory of CENPALAB. The levels of total protein, glucose, alkaline phosphatase, alanine aminotransferase, aspartate aminotransferase, albumin, creatinine, uric acid, and urea were measured a Cobas Integra 400 PLUS Analyzer (Roche DiagnosticSystems).
Vaccination
Four experimental groups (G2-2, G2-3, G3-2, and G3-3) were formed with complete litters comprising 9 and 12 piglets per group. Two groups included 2-week old piglets, and the other two were formed with 3-week old piglets on the day of prime vaccination. Each group was further subdivided to receive the vaccine with either two or three-week intervals between the priming and the booster. The intramuscular route inoculated a dose of 2 mL (50 μg of E2-CD154). The first immunization was performed on the right side of the neck and the second on the left side, using 18 x 1’’ needles, in agreement with the good veterinary clinical practices.
CSFV-neutralizing antibodies detection
The serum samples were tested by Neutralization Peroxidase Linked Assay (NPLA) to determine the NAb titers against CSFV, following the methods described by OIE Manual 16,17. National Center provided Margarita CSFV strain for Animal and Plant Health, Mayabeque, Cuba, and the anti E2 Mab CBSSE2.3 conjugated to horseradish peroxidase by CIGB-Sancti Spiritus, Cuba.
Data analysis
Data analysis was performed using package GraphPad Prism 6 (Prism 6 for
Windows, Version 6.01, GraphPad Software, Inc., La Jolla, USA). The Kolmogorov-Smirnov test was used to evaluate normality, and the Levene test assessed the homogeneity of variances in the data. One-way ANOVA, followed by Tukey multiple comparison tests, was applied to compare the temperature values among groups. Kruskal-Wallis non-parametric test, followed by the Dunn test, was used to compare the geometric means (GM) of the antibody titers among the different groups. Statistical significance was considered when p <0.05.
RESULTS
Evaluation of the clinical signs during the 6 months follow-up
The animals exhibited good health conditions at the beginning of the trial. After vaccination, they showed no signs of inflammation, redness, or induration at the inoculation site. No systemic adverse effects or other alterations were observed, regardless of the age of the animals at vaccination and the vaccination schedule applied.
Evaluation of the body temperature during the immunization period
The rectal temperature was monitored before the immunizations as a part of the assessment of the health status of the animals. Subsequent measures of the body temperature were done at 72 h after each immunization (Figure 1). Mean body temperature in all the groups remained within the physiological range, although some hyperthermia values were observed in G2-2 and G2-3 before vaccination and 72 h later. No significant differences were found between the immunization days and 72 h later; therefore, the high values of temperature registered for some individuals were not associated with vaccination. Animals remained healthy until the end of the trial.
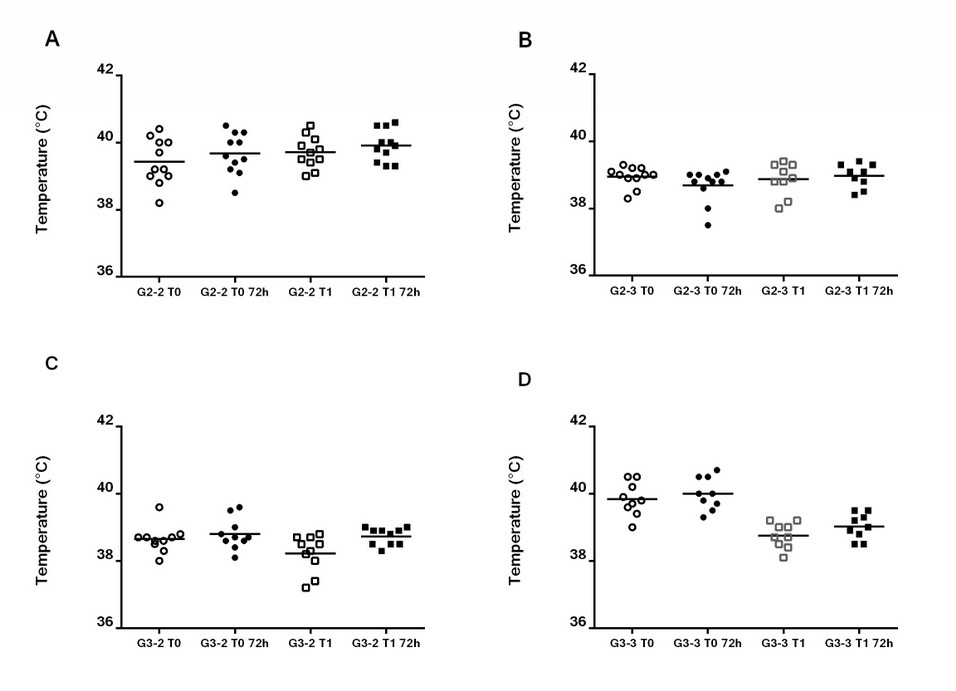
Figure 1. Evaluation of the rectal temperature before and after the vaccination.
Evaluation of hematological and biochemical parameters
The hematological and biochemical parameters of the animals were evaluated before and three months after vaccination (tables 1 and 2). Although some variability was observed at the initial measurements at 15 or 21 days of age, no differences were found among the four groups three months after vaccination (ANOVA p < 0.05). At that time all the values were within the normal ranges for the species.
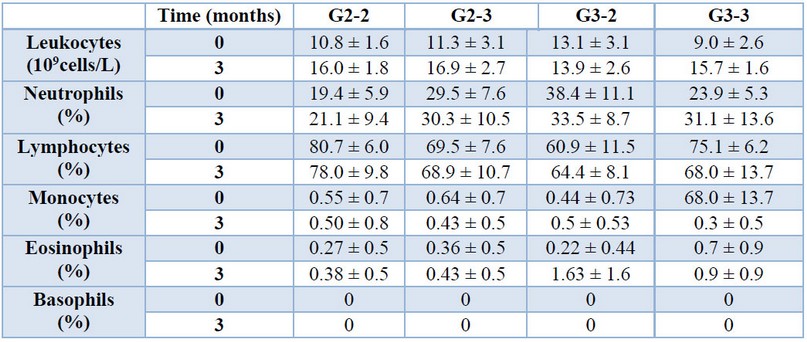
Table 1. Hematological parameters of the animals before and 3 months after vaccination
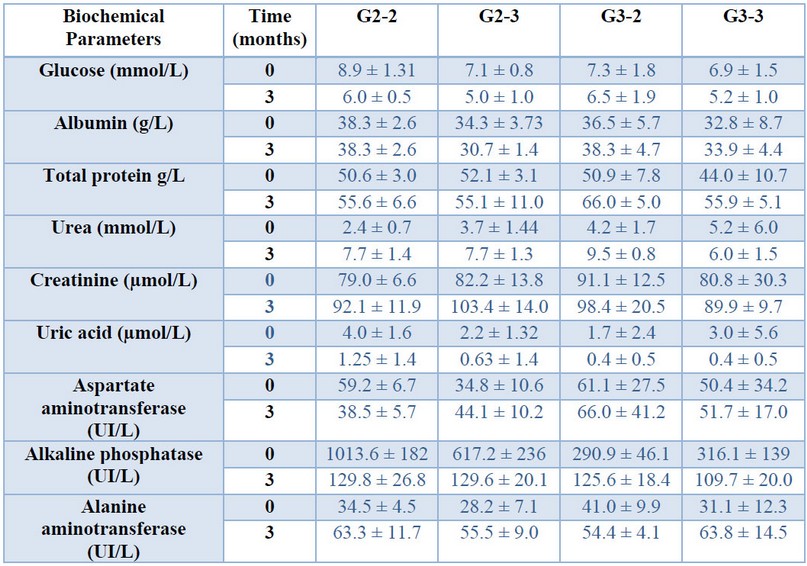
Table 2.Biochemical parameters of the animals before and 3 months after vaccination
Immunogenicity of PorvacÒ in piglets
All piglets were CSFV seronegative at the beginning of the study, with NAb titers lower than 1/5. One month after receiving two immunizations, all animals developed NAb titers of 1/3200 or higher, regardless of the schedule applied (Figure 2). Statistical differences were documented among groups (Kruskal-Wallis, p<0.05). NAb titers were lower in group G2-2 (GM =1/5552) as compared with groups G2-3 (GM = 1/12800) and G3-3 (GM =1/8300) (Dunn test, p<0.05). No significant differences were found between the groups immunized at two and three weeks (G2.2 vs. G3.2 (GM=1/11943) and G2.3 vs. G3.3).

Figure 2. CSFV NAb titers in all experimental groups during the six months follow-up study.
In the evaluation conducted three months after the second immunization, all groups exhibited GM NAb titers above 1/9000, without significant differences among them (Kruskal-Wallis, p>0.05).
Six months after vaccination, the NAb titers had decreased as expected, although they remained equal to or higher than 1/400 in all animals. Finally, six randomly selected animals from the different schemes were followed up one year after vaccination. Interestingly, NAb titers of these pigs remained ≥ 1/400 at this time.
DISCUSSION
Vaccination is the most important activity within the CSF prevention program in endemic countries like Cuba since different pig farms use diverse production systems, the evaluation of different vaccination regimes with PorvacÒ is an important step to adjust vaccination to those production systems.
In this study, we have evaluated the safety and immunogenicity of four immunization regimens with PorvacÒ. Two variables were explored: (1) age for the first administration and (2) time interval between the priming and booster doses.
The four regimens were safe and well-tolerated. Neither systemic nor local adverse reactions were observed nor temperature changes associated with vaccination. Those results confirm pre-registration studies in pigs of different ages with this vaccine10-12,18. They are also by those obtained in a pre-registration study with a CP7_E2alf marker vaccine and animals of similar age 19.
Concerning some elevated temperature values registered during the immunization period, the physiological temperature in pigs ranges between 38.7 and 39.7 ° C, although this range could reach 40.2 °C in piglets during the first weeks after birth 20. Our study was conducted between spring and summer with maximum values of ambient temperature above 30° C. This environmental situation can lead to a condition called heat stress, which is characterized by modifying the physiological constants of the animals 21. Therefore, the experimental manipulations in one of the most sensible pig’s categories, and the heat stress could have influenced the temperature ranges observed.
The development of the innate immune system in pigs begins early during gestation. The newborn piglet already has a naive adaptive immune system with low blood levels of natural antibodies, which recognize the most common pathogens 22,23. Piglets achieve full maturity in their immune system after four weeks of life, which guarantees an adequate immune response to vaccines24. Previous studies conducted with other vaccines have found that five-week-old piglets developed a superior NAb response compared to those immunized two weeks earlier 24.
However, in the present study, even two-week-old naïve piglets could induce a potent NAb response against CSFV after vaccination with PorvacÒ. Similar levels of NAb were measured in those animals, at any of the time points evaluated, compared with the group immunized at the age of three weeks.
Additionally, the findings suggest that a three-week interval between immunizations is more effective than two weeks to induce NAb, although these results were found only one month after vaccination. Those differences faded over time and were found neither at 3 nor at 6 months after vaccination.
Even so, it should be noted that, from a practical point of view, the differences above are irrelevant since the NAb titers elicited by the four regimes studied were manifold higher than the protective threshold of 1/50, previously defined by other investigators 25,26.
The high immunogenicity of PorvacÒ, even at this very early age, is probably due to the action of the CD154 protein (CD40L), which functions as a molecular adjuvant. This molecule is directly involved in the activation and maturation of B lymphocytes, which occupy a central role in the immune response, functioning as precursors of antibody-secreting cells and effective antigen-presenting cells (APC). CD154 also interacts with professional APC such as dendritic cells, promoting their maturation and activation 27,28.
Finally, those results confirm previous findings that PorvacÒ can promote a long-lasting response, which is welcome for breeding gilts and boars, although immune protection for 7–8 months is sufficient for growing and fattening pigs 29. In the present study, when the vaccinated animals arrived at reproductive age still exhibited protective NAb levels against CSFV, demonstrating that a long-lasting humoral immune response can also be generated by immunizing two-week-old naïve piglets PorvacÒ. This agrees with the results of pre-registration controlled trials conducted by Suárez et al., 2011 11, where protective NAb titers were still detected at 9 months post-vaccination.
This study was conducted in naïve animals born to unvaccinated sows; therefore, they did not have MDNA at vaccination. It is well known that MDNA can interfere with active immunity, mainly described for MLVs 24,30,31. However, previous studies have already demonstrated that pre-existing MDNA does not interfere with the immunogenicity of PorvacÒ 18. Therefore, it is very likely that the results shown here could be safely extrapolated to those piglets born to PorvacÒ vaccinated sows.
CONCLUSIONS
A three-week interval between doses induced higher NAb titers than a 2-week interval in piglets vaccinated with 2 weeks of age. Those differences were only observed at 1 month, but not at 3 and 6 months after vaccination.
Regarding the time of the prime vaccination, no differences in the NAb response were found between 2 and 3 weeks of age.
The four immunization schedules evaluated in this study induced high titers of NAbs in naïve piglets, well above the protective threshold; therefore any of them could be used in pig production units, harmonized with their respective productive systems. As a general recommendation, all producers could apply the first dose of PorvacÒ at two weeks of age. The animals born in the genetic farms and reproductive centers would receive the second dose two weeks later. The rest of the farms, including medium and small-scale producers, may apply the second dose with either two or three week’s interval depending on their production systems.
Acknowledgments
This research was supported by the Center for Genetic Engineering and Biotechnology, Havana, Cuba. The author acknowledges all researchers and technicians from the CENPALAB Center for their unconditional support in handling and care of the animals.
REFERENCES
1. OIE. Classical Swine Fever (Infection with Classical Swine Fever Virus).
2. Fonseca-Rodríguez O, Centelles Garcia Y, Alfonso Zamora P, et al. Classical Swine Fever in a Cuban Zone Intended for Eradication: Spatiotemporal Clustering and Risk Factors. Frontiers in veterinary science 2020;7:38.
3. Perez LJ, Diaz de Arce H, Perera CL, et al. Positive selection pressure on the B/C domains of the E2-gene of classical swine fever virus in endemic areas under C-strain vaccination. Infection, genetics and evolution : journal of molecular epidemiology and evolutionary genetics in infectious diseases 2012;12:1405-12.
4. Coronado L, Rios L, Frías MT, et al. Positive selection pressure on E2 protein of classical swine fever virus drives variations in virulence, pathogenesis and antigenicity: Implication for epidemiological surveillance in endemic areas. Transboundary and emerging diseases 2019;66:2362-82.
5. Van Oirschot J. Vaccinology of classical swine fever: from lab to field. Veterinary microbiology 2003;96:367-84.
6. Dong XN, Chen YH. Marker vaccine strategies and candidate CSFV marker vaccines. Vaccine 2007;25:205-30.
7. Bouma A, De Smit A, De Jong M, De Kluijver E, Moormann R. Determination of the onset of the herd-immunity induced by the E2 sub-unit vaccine against classical swine fever virus. Vaccine 2000;18:1374-81.
8. Uttenthal A, Le Potier MF, Romero L, De Mia GM, Floegel-Niesmann G. Classical swine fever (CSF) marker vaccine. Trial I. Challenge studies in weaner pigs. Vet Microbiol 2001;83:85-106.
9. Sordo-Puga Y, Suárez-Pedroso M, Naranjo-Valdéz P, et al. Porvac® Subunit Vaccine E2-CD154 Induces Remarkable Rapid Protection against Classical Swine Fever Virus. Vaccines 2021;9:167.
10. Suárez M, Sordo Y, Prieto Y, et al. A single dose of the novel chimeric subunit vaccine E2-CD154 confers early full protection against classical swine fever virus. Vaccine 2017.
11. Suárez-Pedroso M, Sordo-Puga Y, Sosa-Teste I, et al. Novel chimeric E2CD154 subunit vaccine is safe and confers long lasting protection against classical swine fever virus. Veterinary Immunology and Immunopathology 2021;234:110222.
12. Muñoz-González S, Sordo Y, Pérez-Simó M, et al. Efficacy of E2 glycoprotein fused to porcine CD154 as a novel chimeric subunit vaccine to prevent classical swine fever virus vertical transmission in pregnant sows. Veterinary microbiology 2017;205:110-6.
13. VICH. Good clinical practices, GL09. 2000.
14. VICH. Target Animal Safety for Veterinary live and inactivated Vaccines, GL44. 2010.
15. VICH. Harmonization of criteria to waive target animal batch safety testing (TABST) for inactivated vaccines for veterinary use, GL50 2014.
16. OIE. OIE Terrestrial Manual. Chapter 3.8.3. Classical swine fever (hog cholera) infection with classical swine fever virus. In: OIE, ed.: OIE; 2019.
17. Terpstra C, Bloemraad M, Gielkens AL. The neutralizing peroxidase-linked assay for detection of antibody against swine fever virus. Vet Microbiol 1984;9:113-20.
18. Sordo-Puga Y, Pérez-Pérez D, Montero-Espinosa C, et al. Immunogenicity of E2CD154 Subunit Vaccine Candidate against Classical Swine Fever in Piglets with Different Levels of Maternally Derived Antibodies. Vaccines 2021;9:7.
19. Farsang A, Lévai R, Barna T, et al. Pre-registration efficacy study of a novel marker vaccine against classical swine fever on maternally derived antibody positive (MDA+) target animals. Biologicals : journal of the International Association of Biological Standardization 2017;45:85-92.
20. Mount L, Rowell J. Body size, body temperature and age in relation to the metabolic rate of the pig in the first five weeks after birth. The Journal of physiology 1960;154:408.
21. Carroll J, Burdick N, Chase Jr C, Coleman S, Spiers D. Influence of environmental temperature on the physiological, endocrine, and immune responses in livestock exposed to a provocative immune challenge. Domestic Animal Endocrinology 2012;43:146-53.
22. Butler J, Lager K, Splichal I, et al. The piglet as a model for B cell and immune system development. Veterinary immunology and immunopathology 2009;128:147-70.
23. Lim SI, Song JY, Kim J, et al. Safety of classical swine fever virus vaccine strain LOM in pregnant sows and their offspring. Vaccine 2016;34:2021-6.
24. Suradhat S, Damrongwatanapokin S, Thanawongnuwech R. Factors critical for successful vaccination against classical swine fever in endemic areas. Vet Microbiol 2007;119:1-9.
25. Terpstra C, Wensvoort G. The protective value of vaccine-induced neutralising antibody titres in swine fever. Vet Microbiol 1988;16:123-8.
26. Biront P, Leunen J, Vandeputte J. Inhibition of virus replication in the tonsils of pigs previously vaccinated with a Chinese strain vaccine and challenged oronasally with a virulent strain of classical swine fever virus. Veterinary microbiology 1987;14:105-13.
27. Elgueta R, Benson MJ, de Vries VC, Wasiuk A, Guo Y, Noelle RJ. Molecular mechanism and function of CD40/CD40L engagement in the immune system. Immunol Rev 2009;229:152-72.
28. Xu Y, Song G. The role of CD40-CD154 interaction in cell immunoregulation. Journal of biomedical science 2004;11:426-38.
29. Hua RH, Huo H, Li YN, et al. Generation and efficacy evaluation of recombinant classical swine fever virus E2 glycoprotein expressed in stable transgenic mammalian cell line. PLoS One 2014;9:e106891.
30. Suradhat S, Damrongwatanapokin S. The influence of maternal immunity on the efficacy of a classical swine fever vaccine against classical swine fever virus, genogroup 2.2, infection. Veterinary microbiology 2003;92:187-94.
31. Vandeputte J, Too HL, Ng FK, Chen C, Chai KK, Liao GA. Adsorption of colostral antibodies against classical swine fever, persistence of maternal antibodies, and effect on response to vaccination in baby pigs. American journal of veterinary research 2001;62:1805-11.
Received: 03 April 2021
Accepted: 2 June 2021
Aymé Oliva-Cárdenas1, Fé Fernández-Zamora 2, Elaine Santana-Rodríguez1, Yusmel Sordo-Puga1, Milagros de la C. Vargas-Hernández1, María P. Rodríguez-Moltó1, Danny Pérez-Pérez1, Talia Sardina-González1,Carlos A. Duarte1, Avelina León-Goñi 2, Diurys Blanco -Gámez2, Francisco Contreras-Pérez2, Odalys Valdés-Faure2, Rosmery Hernández-Prado1, Eric Acosta-Lago2, Ileana Sosa-Testé2, Marisela F. Suárez-Pedroso1
Authors Affiliation
1Departamento de Biotecnología Animal, Centro de Ingeniería Genética y Biotecnología (CIGB), Apdo 6162, Playa, La Habana 10600, Cuba.
2Centro para la Producción de Animales de Laboratorio (CENPALAB), Centro de Toxicología Experimental (CETEX), Carretera Tirabeque, Reparto La Unión, Boyeros, La Habana, Cuba.
Corresponding author
Aymé Oliva Cárdenas. Apdo 6162, Playa, La Habana 10600, Cuba. Phone: 53-7-2504421, Fax: (53-7) 271 4764. Email: [email protected]