2022.07.02.2
Files > Volume 7 > Vol 7 No 2 2022
Cost Analysis of Cacao (Theobroma cacao L.) Plant Propagation through the Somatic Embryogenesis Method

Ana María Henao Ramírez 1,* , David Hernando Palacio Hajduk 2 and Aura Inés Urrea Trujillo 3
, David Hernando Palacio Hajduk 2 and Aura Inés Urrea Trujillo 3
1 Center of Agrobiotechnological Development and Innovation – CEDAIT, Universidad de Antioquia, Km. 1.7 vía San Antonio de Pereira - Carmen de Viboral, A.A 054048, Colombia; [email protected]
2 Center of Agrobiotechnological Development and Innovation – CEDAIT, Universidad de Antioquia, Km. 1.7 vía San Antonio de Pereira - Carmen de Viboral, A.A 054048, Colombia; [email protected]
3 Biology Institute, Universidad de Antioquia, Calle 70 No. 52-21, Medellín, A. A 050010, Colombia; [email protected]
* Correspondence: [email protected]; Tel.: +57 3013920674
Available from: http://dx.doi.org/10.21931/RB/2022.07.02.2
ABSTRACT
In vitro cacao (Theobroma cacao L.) production via somatic embryogenesis (SE) is being implemented to mass propagate clonal plant material with the donor material’s prominent characteristics. Though it is an advanced technology, it is con-sidered expensive compared to other propagation techniques. This work focused on identifying the critical financial feasi-bility factors for the SE productive process. The process's costs were estimated, identifying factors influencing each la-boratory's standardized ES process. A Monte Carlo Simulation (MCS) was performed to evaluate different variables upon increasing productive scale in a biofactory (commercial-scale production). The projected lot volume was 100,000 plantlets in solid media, considering the process flow from in vitro introduction to acclimation. A biofactory operational model was projected, establishing time and operator movements and identifying direct and indirect costs.
Costs were defined by the standardized or integral method, with estimated and budgeted calculations to set the cost per plantlet. The identified cost components were culture medium (CM), indirect manufacturing costs (IMC), labor (direct and indirect) and operating expenses. Labor had the most significant share of the costs, at 53%, followed by operating expenses, at 30%, CM, at 12%, and IMC, at 5%. The MCS helped define that the variables with the highest impact on unit price were the embryos’ response in the germination-acclimation stage and the proliferation coefficient during the maturation stage. This projection yielded a figure of US $0.73 per plantlet. However, strategies to reduce this cost have been proposed. These strategies are mainly conducive to optimizing labor and implementing practices that increase multiplication.
Keywords. Plant Tissue culture, Cost analysis, Large-scale production, Biofactory, Monte Carlo Simulation (MCS)
INTRODUCTION
Cacao (Theobroma cacao L.) production as a raw material for the chocolate industry has grown in many countries worldwide. A concentration can be observed in tropical countries, led by African countries, such as Ivory Coast, Ghana, Cameroon and Nigeria, with 63.2% global production 1. In Indonesia and Papua New Guinea, the continent of Asia holds 17.4% of all production, and Latin America, in the countries of Brazil, Ecuador, Peru, the Dominican Republic, and Co-lombia, contributes 19.4% of the global output 2,3. Colombia is the ninth-largest producer worldwide, with a minimum participation of 1.3% despite having significant production potential 4. Colombia’s geographic characteristics, such as its topography, climate, hydrography, and relative humidity, are ideal for developing this crop. These conditions can be found in a significant percentage of the territory 5. Production is currently performed in 29 of the country’s 32 departments, in which Santander, Antioquia, Arauca, Huila, and Tolima stand out, representing 70% of national production 6.
Developing mass propagation systems that facilitate plants' high availability to establish new crops and renew existing old crops is required to increase national production competitively. Asexual or vegetative propagation plays an essential role in reliably reproducing desirable characteristics, and, in the case of vegetative propagation, it is traditionally performed through grafting and root cuttings. However, both techniques have low production levels, the reason why extensive clonal gardens are required to produce enough material, limiting automation and scaling. Moreover, due to genetic improvement programs performed in several research centers worldwide for cacao, there are a considerable amount of improved genotypes 7–10. Nevertheless, one of the largest limiting factors to harnessing this germplasm is the lack of mass cloning methods for the selected financially and agriculturally efficient plants.
In this sense, developing in vitro plant tissue culture techniques allows obtaining plants from one same tissue by taking advantage of plant cells’ totipotence 11. The main advantage of in vitro cultures as a propagation method is the uniform production of plants that conserve the original plant’s characteristics. Besides, the processes are amenable to being scaled up to an industrial level 12. In addition, it allows mass propagating elite plant material in any season, independent of natural environmental conditions, conserving the plants’ genetic potential 13. There is a possibility of obtaining pathogen-free plants (fungi, bacteria, viruses) 14. This technology facilitates optimizing the use of controlled environmental and nutritional factors, optimizing culture areas with large numbers of plants per unit of area, and conserving genotypes selected for prolonged periods. Furthermore, genetically modified plants can be obtained using in vitro regeneration technology, shortening plant breeding program times.
There are various in vitro asexual propagation methods. Among them is multiplication via SE, which, by Mendéz et al. 2019 15, is defined as a process in which a bipolar structure with radial and apical axes similar to a zygotic embryo is developed from a somatic cell without a vascular connection to the original tissue and is capable of growing and creating typical plants. Studies on SE in cacao have been performed for over 40 years 16. Esan 1977 17 was the first to report the formation of somatic embryos, and, since then, numerous studies have been performed to optimize the number of embryos formed by explant 18–21. Subsequently, the work performed by Maximova et al. 2002 22 and Fontanel et al. 2002 23 stands out, in which secondary embryogenesis was proven to allow obtaining a more significant number of regenerants synchronously. In the last five years, the work performed by Garcia et al. 2018 24 has stood out, which reports efficiently obtaining plants via direct SE and in liquid and solid media.
On the other hand, it has been demonstrated that cacao plants resulting from this propagation system have an agronomic behavior without significant differences concerning plants propagated by conventional methods, such as grafting 25–27. Companies, such as Nestlé, have currently standardized the process to establish their crops in countries such as Ecuador, Indonesia, Puerto Rico, Brazil, Ghana, and Ivory Coast 28. There are no reports of large-scale cacao plant material production in Colombia by biotechnological methods, such as SE. SE propagation results have been obtained in solid media for universal genotypes CCN51, TSH565, EET8, ICS1, ICS39, ICS60, ICS95 and IMC67 29 and regional genotypes CNCh12, CNCh13, CNCh16, CNCh24 and CNCh4 30,31.
The financial analysis's important to ensure the production process's feasibility via SE has been mentioned in various studies. It is currently recognized that one of its most challenging aspects is reducing the process's cost 32–34. There has been increased interest in the problems related to large-scale plant production in the last two decades 35–38 and in reducing commercial production costs. However, the accurate details of calculating costs of producing plantlets by SE are seldom discussed or reported 12,39–41. A series of cost reduction strategies have been developed to overcome this limitation in the entire production process, but the cost would largely depend on the effectiveness and efficiency of the SE, which depends on the cacao genotype of interest. It usually has been argued that high propagated material costs, specifically by SE, can be justified by increasing the crop's productivity and uniformity 42. Even though this is plausible, it will not always be achieved until they can prove it at the corresponding production scale and know how much of a return on investment can be obtained and, even more importantly, when it will be obtained 38.
The time factor and frequency of productive lots are essential considerations for a ligneous species like cacao since it requires a long time to finish the entire cycle. In addition, the species is recalcitrant to in vitro cultures and has different responses depending on the genotype 43. Various researchers have estimated the cost of producing plants propagated via SE for lig-neous species and have observed that approximately over 50% of the total cost per plantlet corresponds to labor, clarifying that they are studies performed in developed economies, such as the United States 44–46. Labor is highly competitive in countries like Colombia, circling the US $229 as of October 2020 47, representing an opportunity to develop these technologies.
Nowadays, risk management is an integral part of evaluating any project. Therefore, techniques, such as the Monte Carlo Simulation (MCS), a computerized mathematical approach that allows taking risk into account in quantitative analyses and decision-making, become relevant 48. The study considers repeating a process that generates many random samples linked to specific variables of interest n times. Professionals from fields as disparate as finance, project management, energy, manufacturing, engineering, research and development, insurance, oil and gas, transportation, and the environment using this technique 49. The MCS offers the responsible person or organization the option to make decisions from a series of possible results and the probability that they will occur according to the measures taken. It shows extreme possibilities, the results of taking the riskiest and most conservative estimates, and all the possible consequences of intermediate decisions, which are pertinent for this case study because production on an industrial scale is being discussed.
In this context, and with the Laboratory of Plant Physiology and Tissue Culture of Universidad de Antioquia’s experience with SE in cacao, this study's objective is to analyze the productive process of cacao plantlets by SE from a financial perspective. The process's costs were estimated to identify each standardized stage in the laboratory, and a projection of the production process was performed on an industrial production scale-like biofactory (commercial laboratories used for propagation). The analysis allows planning a pilot for biofactory installations, defining and estimating the factors that in-fluence the cost per obtained plant the most and the least through an MCS. This information helps identify low-cost strategies to apply during technological development.
MATERIALS AND METHODS
Location
The study was carried out on the cacao plants’ productive process via standardized SE at the Laboratory of Plant Physiology and Tissue Culture of Universidad de Antioquia, located in the city of Medellín, in the department of Antioquia - Colombia. The work team has a significant track record researching this topic from 2008 to 13,16,30,31,50,51. The infrastructure of the Universidad de Antioquia biofactory was used to project costs on an industrial scale, located in the municipality of Carmen de Viboral in the department of Antioquia, Colombia. Universidad de Antioquia’s biofactory is the only installation specializing in vitro plant material in Colombia. It has a strategic location 20 minutes from José María Córdoba interna-tional airport. It is a 1,227 m2 building capable of producing up to 7 million plants each year via organogenesis and SE. The installations allow automating and scaling propagation protocols and have an architectural design that harnesses sunlight efficiently all day in growth chambers, a natural water source according to the regional environmental authority and its water purification system. They also have a modular design that facilitates expanding specific areas, an automated 276 m2 greenhouse for plantlet acclimation, and 1,101 m2 of manual operation greenhouses. These structural characteristics allow efficiently using sunlight and water at a low cost. Other authors have reported these factors to be significant within process costs 52.
Stages of Cacao Plant Production via SE
The productive concepts proposed by Egertsdotter et al. 2019 38 for producing ligneous plants were adapted to define the stages of cacao plant production by SE.
Cost Components
The systems engineering methodology for in vitro tissue cultures proposed by Chen 2016 53 was used to identify and select the factors that influence the production process. Three main components were included in the cost structure: direct and indirect labor, the cost of materials and supplies required for plant propagation, which extends to some indirect manufac-turing costs, and, lastly, operating expenses, which include the infrastructure, depreciation, and administrative expenses required to launch the business. All parameters and their composition are listed in detail in supplement 1.
Assumptions
The following premises were considered to project cacao propagation costs:
• Losses due to contamination and necrosis depend on the quality of the initial tissue and the planting process.
• Multiplication coefficients and percentage of explants' response depend on controlling subculture times and environ-mental factors, such as light, temperature, and relative humidity.
• Work performed by employees is calculated under-skilled labor's operational efficiencies with at least one year of ex-perience handling explants in each stage.
• Labor standards can change depending on the hired technical personnel’s training time and experience.
• Thirty percent of the total is regarded to cover the biofactory’s administrative expenses, in which operating expenses and utility are considered.
• The practical exercise was performed to produce 100,000 plants, equal to 1.4% of the biofactory’s installed capacity.
• The performed analyses are a projection of data obtained in the laboratory over different years. They allow establishing a baseline of work by simulating productive factors.
The analysis was performed based on the whole plant produced by lot to begin identifying cost components, considering the number of explants, vessels, and quantity of culture media for each process stage through the following variables and formulas listed in table 1.
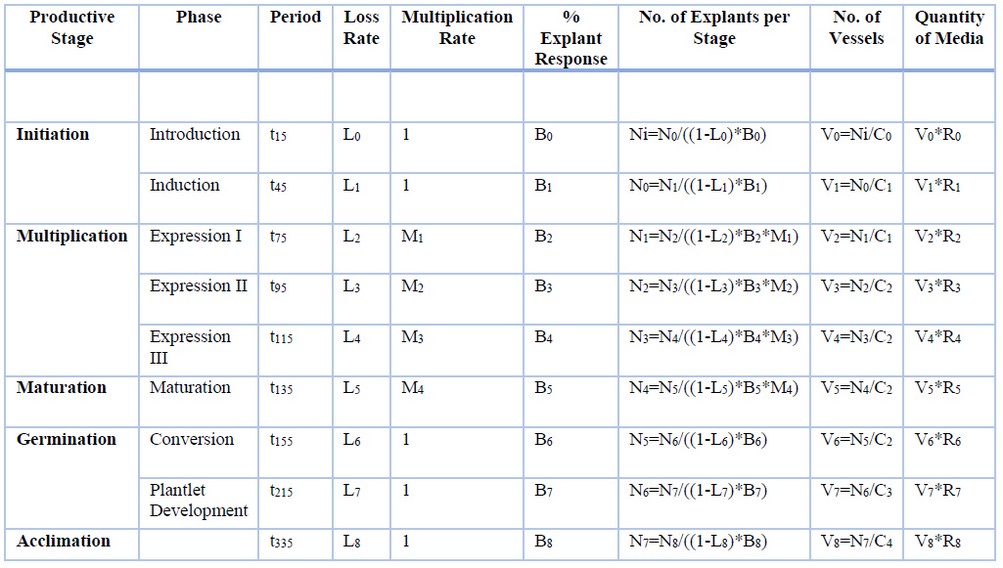
t (Time): The period each stage of the production process requires, measured in days; L (Loss rate): Measured as a percentage, this is each productive stage’s losses due to contamination or necrosis; M (Multiplication rate): The multiplication coefficient of plant material during the multiplication and maturation stages. It is produced at different moments during embryo development. Five times the initial number was obtained in M1 and M2, and it doubled in M3 and M4 (ten); B (Explant response): Measured as a percentage, it is equal to the rate of explant response in each stage; N (No. of explants per stage): This is the number of explants processed in each production stage. The model projects labor requirements and materials in accordance with the desired plants at the end of the process. For this reason, N8 was calculated as the number of cacao plants desired at the end of the process, N8 = 100,000. It is added, for the calculation, that Ni (initial) represents the flower buds entering the process, which have 5 staminodes, for which reason it increases by 5 from Ni to N0; V (No. of Vessels): This is the number of vessels required in each production stage. The calculation is performed based on the number of processed explants and between each type of vessel’s capacity; C (Vessel Capacity): This refers to the number of explants in accordance with the type of vessel used in each stage. The vessels are conical tubes, Petri dishes, magenta vessels and germination trays. The assigned variables are: C0 is a conical tube with a capacity for 350 flower buds, C1 is a Petri dish with a capacity for 25 calli, C2 is a Petri dish with a capacity for 49 embryos, C3 is a magenta vessel with a capacity for 20 plantlets, and C4 represents trays with a capacity for 50 plantlets; R (Culture medium): Equal to the volume of culture media required by the type of vessel in a 50 ml conical tube, 30 ml Petri dish, 150 ml magenta vessel and 100 g germination trays; Q (Quantity of Media): This is the result of the number of vessels required in each productive stage by the necessary media volume per vessel.
Table 1. Variables and formulas for calculating the number of explants, number of vessels and quantity of culture media by SE productive stage.
Culture Medium (CM)
Each productive stage has different culture media formulations following the developmental stage of SE to be induced: induction - INDI, multiplication (an expression I - INDIexp, expression II - CM2, expression III - EM2), maturation EM2, germination - MM6 (Table 1 S1: Composition culture medium), and the cost may differ (Sheet 4 S2: Culture medium). The culture medium is calculated with the formula (1) (Table 2). On the other hand, it is essential to remember that most culture media supplies are imported, and costs are affected by the Representative Market Rate (RMR).
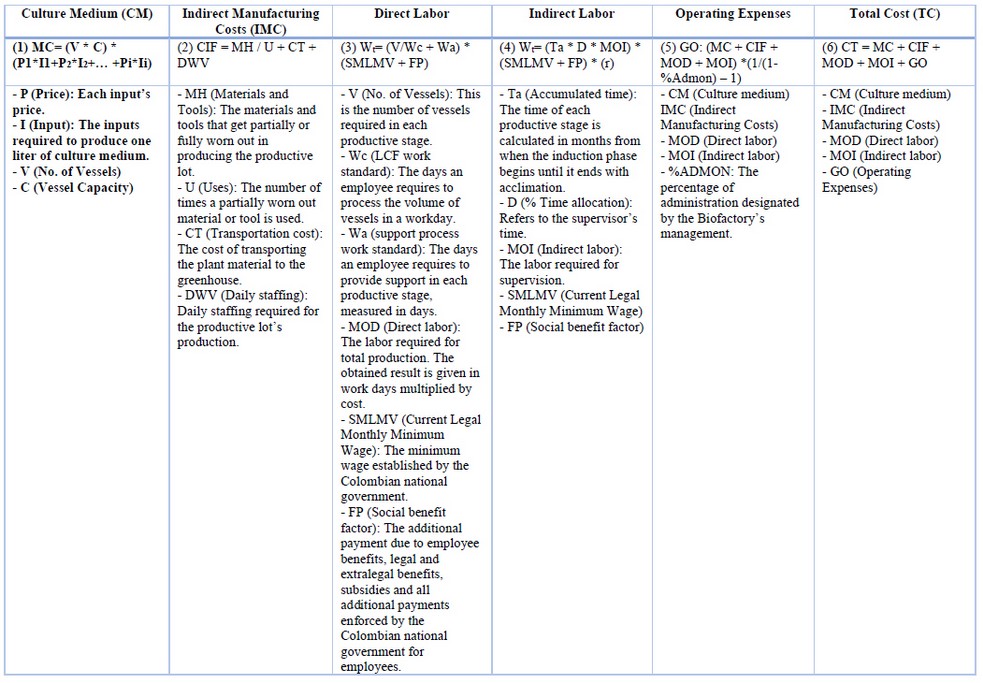
Table 2. Formulas and variables for calculating the components of the productive process’ cost.
Indirect Manufacturing Costs (IMC)
Indirect manufacturing costs are part of the production but cannot be allocated to a production plant or lot because they also have a staggering behavior. Indirect manufacturing costs are calculated using formula (2) (Table 2).
Labor
Direct labor includes the hours required to collect the plant material used as ex-plants (flower buds), which extends to packaging and shipping them. It also in-cludes work in the laminar flow cabinet (LFC) and works performed in support activities, such as preparing, dispensing, and replacing culture media; preparing and disposing of materials; washing vessels; monitoring and selecting.
A supervisor also provides indirect labor with functions related to coordinating direct work, calculating the number of necessary dishes and media, managing chemical products and media stock, equipment, and maintenance for the critical areas. Moreover, they are responsible for ensuring all explants begin production without contamination, on time, and in the correct amounts. They are responsible for maintaining productivity, and, most importantly, they will decide whether or not crop production must be continued, harvested, or discarded at their discretion. The supervisor must coordinate decisions on what product, how many units, and when and where to ship with the administrative department during packaging and shipping.
Direct labor is calculated with the formula (3), and indirect labor is calculated using formula (4) (Table 2).
Operating Expenses
The biofactory’s operating expenses include management, sales, equipment depreciation (caster racks, laminar flow, pH meter, balance, magnetic stirrer, auto-clave, distillation unit, minor Equips, mist chamber, fridge, etc.), leasing and public utilities (water, light, Internet, telephone), among others. A historical average of the biofactory's years was considered to calculate operating expenses. Operating expenses are calculated using formula (5) (Table 2).
Finally, the total costs associated with production are calculated following formula (6) (Table 2).
Monte Carlo Simulation (MCS)
The RMR, loss percentage, percentage of explants with responses, multiplication coefficient, and standard productivity variables are defined in each of the production stages and phases of SE as variables of interest to establish their impact on costs when modifications are made to the productive process. Each one of the variables is assigned a triangular distribution, which is traditionally used for cost analyses. Three model estimation points are defined, placing the laboratory results as the central value, and a 10% variation range is established above and below each variable. A triangle-shaped distribution takes form from the above, where the highest probability is around the middle.
A significant number of simulations can be performed using the MCS analysis, modifying the established ranges for each variable. Having all variables in a similar range allows identifying the most strategic variables on which the team's work should focus to obtain the best results. In this study, the MCS was performed with @risk 8.0.1 54 software, with 100,000 iterations for each variable and 100 simula-tions of the same process. The response objective was cost per plant.
RESULTS
Stages of Plant Production via SE
The production process for the in vitro propagation of cacao plants by SE was identified to have five stages: initiation, multiplication, maturation, germination, and acclimation (Figure 1). Various processes, such as the explant disinfection process, called introduction, and callogenesis induction, are included in the initia-tion stage. In the multiplication stage, undifferentiated tissues are transferred to the primary embryo expression medium with a first multiplication coefficient of 5 for the phase called an expression I. An average of 5 embryos are produced for each callus. Once primary somatic embryos are obtained, they are divided using a blade. The pieces are placed in the repetitive or secondary embryogenesis induction media with a second multiplication coefficient of 5. This phase is called expression II. Therefore, somatic embryos develop asynchronously and are harvested from the globular and early cotyledonary stage to prevent embryonic axes from fusing during development. The embryos are subsequently transferred to a hormone-free medium where repetitive embryogenesis occurs again, with a multiplication coef-ficient of 20. This is called the expression III phase. Up until this point, both the initiation and multiplication stages are performed in a dark room at an average temperature of 26 – 27 ± 2 °C. In the maturation phase, somatic embryos develop with clear differentiation between their apical and radicular meristematic poles. Embryo maturation is set off by a change in the culture medium's composition, where the source of carbon and growth regulators are vital components. Subsequently, the germination stage is composed of the conversion phase to plantlets and their development in terms of increased stem length, secondary root formation, and leaf formation. They are transferred to 500 ml culture vessels when they reach a certain height. Plantlets are kept under natural lighting in growth rooms during the maturation and germination stages. Finally, plantlets are transferred to greenhouse conditions for growth and elongation during the acclimation phase. Plantlets that are 3 - 5 cm high, with 1 - 3 leaves and both primary and secondary roots, are se-lected, removed from their culture vessels, and washed with topwater to eliminate culture medium surpluses. They are then transferred to 50 alveoli germination trays with a substrate made of a mixture of sand and unenriched basic coconut coir. The tray is hermetically sealed for one day, and holes are subsequently opened to allow gas exchange, keeping the substrate hydrated with a Hoagland solution 55. After this time, the tray is opened and kept in the ideal growth conditions for cacao in this stage, with a 50% shade percentage, the temperature of 24 – 30 ºC, and relative humidity over 60%.
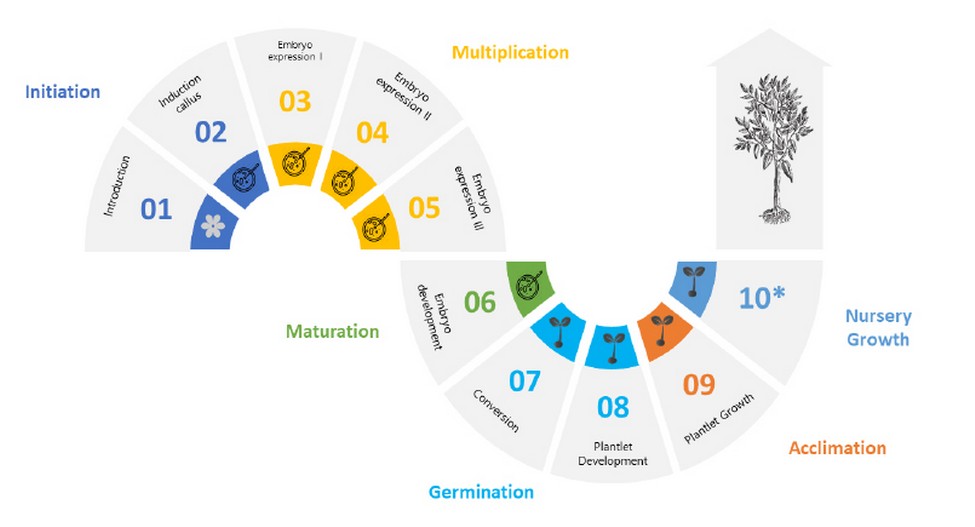
Figure 1. Stages of cacao plant production via somatic embryogenesis (SE).
* Plantlets growth in greenhouses is not considered in this study.
Cost Components
It was determined that Ni 4190 flower buds are required during plant material introduction, and N0 9429 staminodes must be processed to induce callogenesis to produce a batch of 100,000 plantlets (Sheet 2 S2: Stages of production). For the multiplication stage, N1 + N2 + N3, a total of 27,827 embryos were processed. 101,189 embryos were processed in the N4 maturation stage, and 506,803 plantlets were processed in germination stages N5, N6, and N7 (Table 3).

Table 3. Total number of explants processed for each productive stages.
Considering the following amounts and sufficient work times is required for labor payments in Colombia, as presented in the Substantive Labor Code (Sheet 3 S2: Direct and indirect labor). The base salary in Colombia is US $250.8, transportation allowance US $29.4, health US $21.3, pension US $30.1, occupational risks US $1.3, severance payments US $22.6, premium US $23.3, losses US $23.3, interest on severance pay US $2.8, vacation US $10.5 and endowment US $12.5. The sum of the total cost of a salary in Colombia is US $428. To calculate effective work time, it was decided that, of the year’s 365 days, 52 are Sundays, 18 are holidays, 15 are vacation days, and an average of 3 are taken off due to incapacity. Therefore, there are 277 effective workdays a year and 23 effective workdays a month. Fol-lowing the above, taking a salary into account with the social benefit factor and effective monthly workdays, yields US $18.53/effective workday.
Once the daily labor cost was identified, the time personnel used to perform work in each process's phases was calculated. Three main activities were identified: (1) collecting flower buds from the genotypes of interest in the field, (2) processing material in an LFC, and (3) support processes, among which are preparing, dispensing, and replacing culture media, preparing materials, reviewing to discard material, cleaning functions and vessel and tool disinfection (Table 4).
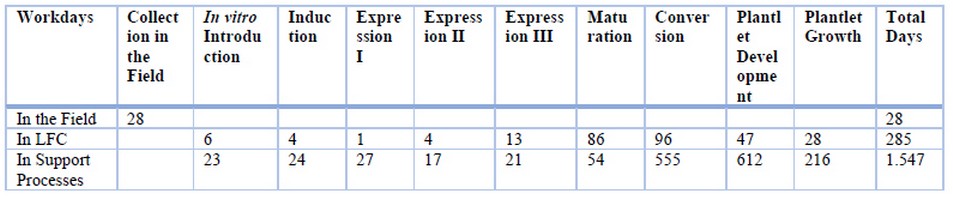
Table 4. Personnel days allotted to propagation activities and support processes in each one of somatic embryogenic phases.
A total of 28 days, or 168 hours of work (approximately), were required to collect flower buds in the field. Personnel working in the LFC involves 285 days or 1,710 hours to execute all the productive process' stages, and support personnel requires 1,547 days of work or 9,286 hours. Therefore, the production process requires 334 days from flows bud introduction to plantlet attainment, and 7 employees are re-quired to execute the process.
The cost of personnel was obtained by multiplying the 1861 required days for batch production by the cost of one effective monthly workday at US $18.53/day, for US $34,500. Supervision does not need to be project-exclusive once standardized processes are achieved, for which reason a 25%-time allotment was established during the production year. A monthly price with a social benefit factor of US $1,284 was obtained for the supervisor, 12 months. Therefore, the total cost of indirect labor was US $3,852.
It was observed that the germination stage contributes significantly to the total cost for culture media since it requires more culture media than other stages (Sheet 4 S2: Culture medium). The amount goes from 30 ml in a Petri dish to 150 ml in a higher-volume vessel (Table 5). For IMC, transportation resulted in US $543, personnel endowment US $282, and materials and tools US $3,049, which adds up to US $3,874. It was previously clarified that operational expenses are calculated as 30% of total production costs. Therefore, they correspond to US $21,895 (Sheet 5 S2: IMC).
Using this estimation methodology, the total cost of US $72,986 was obtained for a production batch with 100,000 plants, with a cost per plantlet of US $0.73/unit.
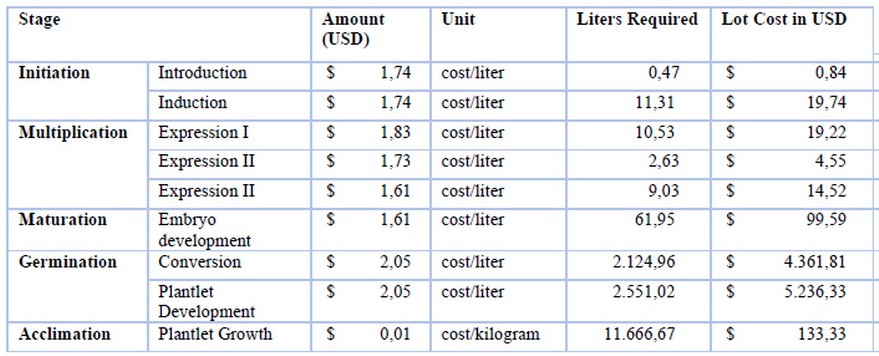
Table 5. Culture media costs in each stage of via somatic embryogenesis (SE) production.
Monte Carlo Simulation Analysis (MCS)
After structuring costs, the most influential cost component was direct labor, representing 53% of the total cost. The cost of culture media was 12% of the total, IMC represented 5%, and operating expenses, including administrative expenses and infrastructure, were 30% (Figure 2).
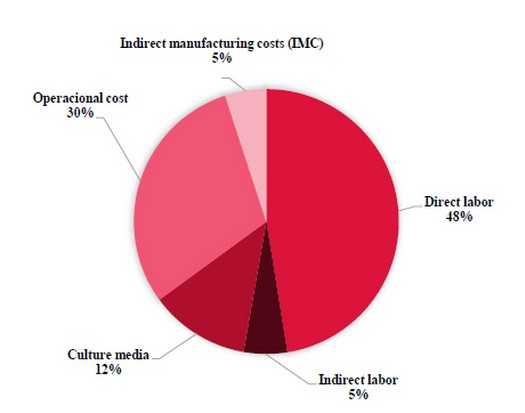
Figure 2. Components of production cost share in the total cost of producing cacao plantlets.
Plants propagated by SE. Figure 3 shows that the cost of plants per unit can be inferred between USD $0.6835 and USD $0.7786, with an average USD $0.7290 (Sheet 1 S2: Total cost), due to the production process's cost structure. If strict control is maintained over the variables while executing the productive lot as es-tablished in this study, the average price per plantlet has been proposed to be USD $0.7290, with a 95% reliability. However, as one may observe, there is a certain asymmetry towards the right, which indicates that the process could increase in cost. That is to say, the cost may have deviated over the average or over USD $0.7290 after moving the lot.
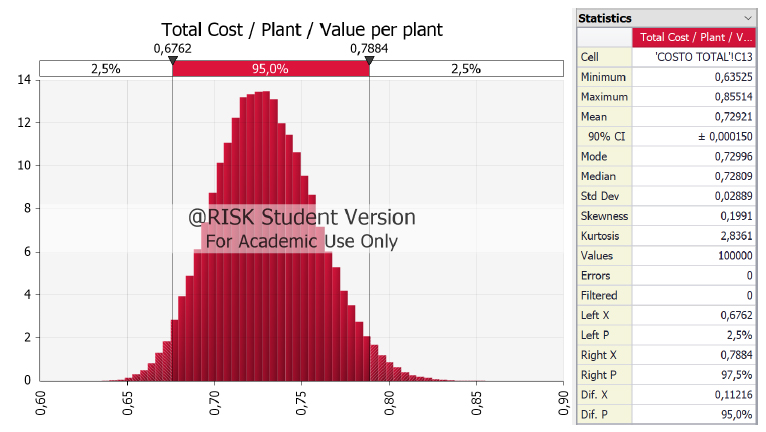
Figure 3. Frequency distribution for the cost of producing one cacao plantlet by somatic embryogenesis.
On the other hand, a regression was performed on the correlation coefficients of each of the cost model’s variables to identify which ones influence the variable response, which is to say cost the most. This analysis showed that the most significant variables are in the productive process’ last stages, corresponding to ger-mination and acclimation, followed by the maturation stage. It was specifically found that the plantlet growth (-0.69) and plantlet development (-0.60) phases, as well as the plantlet conversion phase (0.35) had more significant effects on cost, with 95% reliability. Therefore, when these variables, which are expressed as the percentage of explants' response, rise above 60%, 50%, and 50%, respectively, the variable of cost per plantlet decreases. During the embryos' development phase, both the multiplication coefficient (-0.07) and percentage of explants' response (-0.07) tend to decrease cost per plantlet as their prices increase. They are currently at 10% and 70%, respectively. Besides, it was observed that the RMR (0.15) has a positive effect on cost. Cost per plantlet will increase as RMR increases (Figure 4).
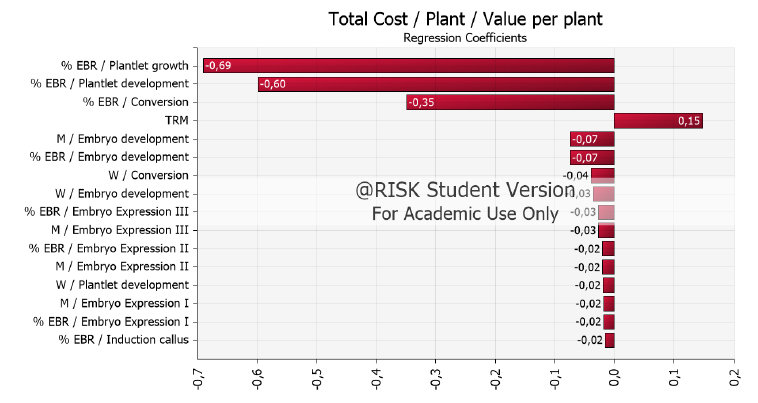
Figure 4. Regression coefficients for the variables associated with the productive process of cacao plants propagated by somatic embryogenesis over the production cost per plant. Variables: % EBR (% explants with response), RMR (Representative Market Rate), M (Multiplication rate), W (Labor standard).
DISCUSSION
In Colombia, approximately 176,000 cultivated hectares benefit 52,000 families in 422 municipalities of 30 departments, with biannual growth of 4% in planted area. It is established that approximately 7000 hectares biannually is required in the short term and some 10,000 hectares for renovation. Having an average amount of 1,100 plants per hectare, with 5% additional plants to replace those that die at the time of being transplanted, it is estimated that the potential demand is 19.6 million plants biannually for 2021 56. In the last two decades, there has been a growing demand for high-quality, high-performance, and pathogen-free planting material, increasing demand for agricultural, forest, and horticul-tural products. Within this trend, cultivating plant tissue in vitro has become an important, commercially feasible tool for generating high-quality, high-performance, and pathogen-free planting material regardless of climate fluctuations. Most notably, SE's potential as a propagation method has not only been described in the last years for cacao 24,57,58 but also other plant species of in-terest 59–61. Various authors agree that the main factor limiting SE's deployment on a commercial scale is the plants' relatively high cost compared to those cultivated in greenhouses 62. Therefore, financial cost analyses for these processes are vital, and more so is examining strategies to reduce costs without affecting production efficiency and quality 41.
In the analysis performed for cacao, the component that contributed most to cost was direct labor, at 53%. This result coincides with what other authors reported for plants obtained by SE other than cacao. For example, Cervelli and Senaratna (1995) 63 reported a 70% contribution to cost for labor and Chu (1995) 64 reported a 63.8% contribution. However, other authors recommend decreasing direct labor's con-tribution percentage as much as possible, by between 30 and 40% 36,52. Following these results, it is evident that cacao plantlet production via SE is a labor-intensive system, and labor is the main production cost component. Maintaining high labor efficiency is essential to make the process feasible. Therefore, one of the method-ological strategies to achieve this objective is optimizing personnel’s actions in LFCs. For example, unnecessary existing times can be eliminated. Despite having adequate efficiencies in processing material, such as the number of transfer opera-tions per day, this study has estimated 6,000 explants/ workday for the initiation stage, a work standard similar to that which Ahloowalia & Savangikar (2004) 36 reported for this same operation, at 5,000 explants/ workday.
On the other hand, following Suárez-Castellá (2006) 65, the productivity of 4,595 explants/ workday can be achieved by providing training on handling and pro-cessing tissue. This figure is higher than the work standards for the cacao multiplication stage, which oscillates at 1,176 explants/ workday on average. However, lack of skilled labor is always the main problem in countries such as Colombia. Implementing training programs is an alternative to promote operating personnel’s work capacity and overcome this limiting factor.
The second component contributing most to cost is operating expenses, at 30% of the total cost. This percentage is very close to the ranges reported when prop-agating other species, such as Phalaenopsis sp., where operating expenses were 32.7% 66. However, lower figures are reported for Paulownia tomentosa produc-tion, at 19% 45 and Saccharum officinarum, 24.4% 67. It must be clarified that both are productive processes performed by organogenesis. In this respect, it is essential to indicate that one of the highest infrastructure costs is electricity associated with air conditioning systems used to regulate culture room temperatures. According to Tomar (2010) 52, this factor can consume 85% of the electricity in vitro propagation installations. Therefore, if the process does not require exact control over temper-ature, it is recommended facilities avoid using this resource if the area's climate conditions allow it. Another aspect of electricity consumption is associated with expenses during the water distillation process, which is the main component of culture media. Distilled, double-distilled or deionized water is generally used in in vitro tissue cultures. However, on a productive scale, according to Sahu (2013) 68, using alternative water sources is recommended to reduce the cost of sterilizing tap water in an autoclave, as long as the water has proper conductivity, pH, is free of heavy metal or pollutants 69.
The third most important component within the cost structure is culture medium, which corresponds to 12% of the total cost and can also be improved to reduce costs. Most plant tissue culture media's main components are mineral salts and sugars as sources of carbon and water. Other components can include organic supplements, growth regulators, and gelling agents 70. According to Prakash et al., (2004) 71, the chemical reagents of culture media cost less than 15% of the total production cost of the vitroplant. In some cases, the cost can lower down to 5%. Of culture media components, gelling agents, such as agar, contribute to 70% of the total cost, followed by sources of carbon and, finally, growth regulators, which minimally influence the cost of production, since they are reasonably inexpensive due to their minimal concentration within culture media 12. Carbon sources are commonly used, such as glucose, fructose, and maltose. However, they are reagents with high purity levels, making them expensive. Sucrose is the most widely used carbon source in the in vitro propagation of plants. Table sugar has also been used as a strategy to reduce culture media costs. Regular sugar has a high enough quality for micropropagation, and using it reduces media culture costs between 78 and 87% 41. In Colombia, the cost of local sugar is USD $0.86 / kg compared to USD $8.13 / kg for imported sucrose. Another strategy for decreasing costs is associated with the mineral formulation. Many companies sell media prepared as a liquid or powder. However, the benefits and disadvantages of formulations prepared in laboratories must be considered concerning commercially prepared media. Although prefabricated media save time, their relative cost is high. Pre-packaged media preparations are usually helpful when the required amounts are small - generally on a scale for research - and they have less errors. It is much more cost-effective for large-scale use to prepare media by combining the basic in-gredients. Nevertheless, if errors occur, losses are more significant depending on the size of the lot. Moreover, the time the hired technician takes preparing the formulations must be considered, for which reason it is recommended for stock solution concentrations to be up to 1,000X.
Indirect manufacturing costs are included in the fourth component, with 5% cor-responding to transportation, culture vessels, materials, supplies, tools, etc. Some authors recommend pre-sterilized, reusable plastic Petri dishes because the material is more inexpensive than glass 71. However, our organization encourages mini-mizing its ecological impact by substituting single-use supplies or tools for lasting and reusable materials due to environmental, social responsibility policies. Fur-thermore, following Datta (2017) 41, using supplies, such as aluminum, is not recommended for wrapping tools before sterilization. Instead, sterilizable stainless steel containers can be used in autoclaves as substitutes with significantly long valuable lives. Also, PVC or polyethylene plastic film is used as lids in common practice and for large-scale production. These lids have been replaced with stainless steel or polypropylene screw caps sterilizable in an autoclave.
The sensitivity analysis showed that the most significant variables on cost per plantlet are in the productive process’ last stages, corresponding to plantlet ger-mination and acclimation. This specifically applies to the percentage’s response during the plantlet conversion, development, and growth stages. These results are consistent given the accumulated time, effort, and energy invested up to this point in the productive process. Following Von Aderkas et al. (2016) 72, practices per-formed during the entire production process via SE, especially managing embry-ogenic structures and somatic embryos in cotyledonary states, significantly affect the performance and quality of the embryo conversion, as well as production costs. One of the most crucial stages is when plantlets are transferred from in vitro to ex vitro conditions since approximately 8 months have transpired in the process’ previous stages. It is well-known that plantlet survival can be improved by optimizing somatic embryos' maturation and germination conditions in cotyledonary states. These conditions can be optimized by, for example, making adjustments to components, such as abscisic acid and gibberellic acid and modifying the culture medium's water potential and nitrogen source, among others 73–75. In this process stage, the production cost can also be reduced if time in hardening facilities is shortened without reducing plantlet survival rates. Plants require relatively more minor time to harden if natural light is previously used during plantlet development 41 and if microorganisms that promote growth, such as fungi and bacteria, are used 76.
The variables associated with the maturation stage, which is to say the development phase of embryos, such as multiplication coefficient and percentage of explants' response, were also significant on cost in the sensitivity analysis. Under other au-thors and the obtained results, improving embryo multiplication rate is one of the most efficient ways to reduce costs 66. Some of the alternatives for improving the multiplication rate in the case of SE include selecting the most suitable develop-ment stage of primary embryos or primary embryonic tissues as a source for pro-ducing repetitive embryos. That is to say, the more control there is over the exact point in which tissues answer to recurring embryo formation, the more embryos will be produced. The same applies to using substances to promote embryo formation, such as demethylating agents like 5-Azacytidine (5-azaC). In this sense, in terms of cacao, it has been demonstrated that decreases in embryogenic potential seem to be related to DNA methylation 77. In addition, environmental conditions like micro-climate, at least while controlling temperature between 27 and 28 °C and total darkness in the culture room, are definitive in the cacao multiplication stage. In this context, all these stages add up cumulatively, reducing production costs.
A price per plantlet of USD $0.73 was obtained based on the analysis performed on the cacao production process, with a final sales price to customers of USD $0.88 and grafted of USD $1.16. Upon reviewing the market prices of traditionally grafted cacao plants, an average of USD $0.71 was obtained for Colombia and, in countries like Ecuador, USD $0.70. Also, the cost in Colombia is above the prices offered in Latin America. For example, cacao plantlets propagated by SE in a biofactory in Brazil cost USD $0.70 78. Despite there being a significant difference concerning plantlets propagated by the traditional method, there is a significant opportunity to be considered by developing the technology on a biofactory scale with obvious cost reduction objectives. In this sense, to validate technology for cacao plant production by SE and for it to be a cost-effective product on the national market, the factors that contribute most to cost, such as percentage of explants response, must continue to be optimized, and other options that do not compromise quality must be considered. Adopting flawed low-cost strategies can make the production process prone to failure since it must be highlighted that this is a productive process inherent to a biological process. These processes may vary significantly after changing some culture conditions. Low-cost techniques will only be successful if they scrupulously comply with tissue culture's primary require-ments, thus maintaining plantlet quality. According to Datta (2017) 41, procedure standardization is more critical for ensuring cultivated plants' quality than proce-dure sophistication. It can be said that continuing to develop a low-cost productive process signifies an advanced generation technology, improving the process’ effi-ciency and use of resources. Therefore, different strategies can be intelligently adopted, such as simplifying various technological operations to reduce specialized facility costs, such as a biofactory’s costs.
CONCLUSIONS
A calculation matrix that estimates production costs by defining the number of plants required by productive batch and estimating cost developing volume was created in this study. Labor costs contributed to 53% of the total cost, followed by operating expenses at 30%, culture media at 12%, and IMC at 5%. The proposed costing and analysis approach can be adapted and applied to other cultures propagated by SE.
The percentage of explants' response variable in the germination and acclimation stages is considered to affect cost per unit most significantly for cacao plantlet production by SE and the multiplication coefficient during the maturation stage.
The applicability of the MCS was proven, showing that it is possible to consider different strategies for reducing costs. In this sense, nowadays, deciding what productivity indicator should be used to consider these scenarios is practiced. It represents an opportunity for developing technology and business. Finally, it is essential to mention that the presented cost reduction strategies must be tested on a short-term pilot batch to validate the product’s performance.
Author Contributions: Conceptualization, Ana María Henao Ramírez and Aura Inés Urrea Trujillo; methodology and software, Hernando David Palacio Hajduck and Ana María Henao Ramírez; validation anf formal analysis, Ana María Henao Ramírez; investigation, resources, data curation, writing—original draft prepara-tion, Ana María Henao Ramírez; writing—review and editing and supervision, Aura Inés Urrea Trujillo. All authors have read and agreed to the published version of the manuscript.
Funding: This research was funded by General Royalties System - Science, Technology, and Innovation Fund with the Center of Agrobiotechnological Development and Innovation– CEDAIT- BPIN 2016000100060, National Planning Department, Office of the Governor of Antioquia, Universidad de Antioquia, Universidad Católica de Oriente and Compañía Nacional de Chocolates.
Acknowledgments: We would like to thank Laboratory of Plant Physiology and Plant Tissue Culture of the Universidad de Antioquia. A special acknowledgment to Universidad de Antioquia’s Research Development Committee (CODI) and Granja Yariguíes – Compañia Nacional de Chocolates.
Conflicts of Interest: The authors declare no conflict of interest.
REFERENCES
1. Toma S, Săseanu A. Chocolate, a Global Business. In: BASIQ 2020 International Conference. ; 2020:1233.
2. ICCO. Quarterly Bulletin of Cocoa Statistics. International Cocoa Organization. Published 2019. Accessed October 16, 2020. https://www.icco.org/about-us/international-cocoa-agreements/cat_view/29-quarterly-bulletin-of-cocoa-statistics.html?limit=35&limitstart=0&order=name&dir=DESC
3. Wickramasuriya A, Dunwell J. Cacao biotechnology: current status and future prospects. Plant Biotechnol J. 2018;16(1):4-17. doi:10.1111/pbi.12848
4. Abbott P, Benjamin T, Burniske G, et al. An Analysis of the Supply Chain of Cacao in Colombia.; 2019. doi:10.13140/RG.2.2.10934.14400
5. Hartemink A. Nutrient stocks,nutrient cycling, and soil changes in cocoa ecosystems. In: Advances in Agronomy. Elsevier; 2005:227-253. doi:10.1016/s0065-2113(05)86005-5
6. Procolombia. The main regions of Colombian cocoa production | Colombia Country Brand. Published 2019. Accessed January 14, 2020. https://www.colombia.co/en/colombia-country/colombia-the-land-where-cocoa-production-is-cacao-fino-de-aroma/
7. Ofori A, Arthur A, Padi FK. Extending the cacao (Theobroma cacao L.) gene pool with underrepresented genotypes: growth and yield traits. Tree Genet Genomes. 2019;15(5):75.
8. Fister AS, Landherr L, Maximova SN, Guiltinan MJ. Transient expression of CRISPR/Cas9 machinery targeting TcNPR3 enhances defense response in Theobroma cacao. Front Plant Sci. 2018;9:268.
9. Duval A, Gezan SA, Mustiga G, et al. Genetic parameters and the impact of off-types for Theobroma cacao L. in a breeding program in Brazil. Front Plant Sci. 2017;8:2059.
10. Casanova TJ, Amores FM, Garzón AI, et al. Costs and temporary distribution of the investment for the genetic improvement and development of a clonal variety of cocoa (Theobroma cacao L.) of high productivity. In: International Symposium on Cocoa Research (ISCR), Lima, Peru, 13-17 November 2017. International Cocoa Organization (ICCO); 2018.
11. Fehér A. Callus, dedifferentiation, totipotency, somatic embryogenesis: What these terms mean in the era of molecular plant biology? Front Plant Sci. 2019;10(April):1-11. doi:10.3389/fpls.2019.00536
12. Thompson D. Challenges for the large-scale propagation of forest trees by somatic embryogenesis - a review. In: Proceedings of the 3rd International Conference of the IUFRO on “Woody Plant Production Integrating Genetic and Vegetative Propagation Technologies.” ; 2014:81-91.
13. Henao-Ramírez AM, Jaime H, Duque S, et al. Determination of Genetic Stability in Cacao Plants (Theobroma cacao L.) Derived from Somatic Embryogenesis Using Microsatellite Molecular Markers (SSR). Int J Fruit Sci. 2021;00(00):1-15. doi:10.1080/15538362.2021.1873219
14. Edward R, Wetten A. Virus Detection and Elimination in Cocoa (Theobroma cacao L.) Through Somatic Embryogenesis. J Plant Sci. 2016;4(3):52. doi:10.11648/j.jps.20160403.14
15. Méndez H, Ledezma M, Avilez R, et al. Signaling overview of plant somatic embryogenesis. Front Plant Sci. 2019;10:1-15. doi:10.3389/fpls.2019.00077
16. Henao-Ramírez A, Urrea-Trujillo A. Somatic Embryogenesis for Clonal Propagation and Associated Molecular Studies in Cacao (Theobroma cacao L.). In: Chong P, Newman D, eds. Agricultural, Forestry and Bioindustry Biotechnology and Biodiscovery. Springer, Cham; 2020:63-102. doi:https://doi.org/10.1007/978-3-030-51358-0_5
17. Esan E. Tissue culture studies on cocoa (Theobroma cacao L.) A supplementation of current research. In: Proceedings V International Cacao Research Conference, Cacao Research Institute of Nigeria. ; 1977:116-125.
18. Ajijah N, Hartati R. Primary and secondary somatic embryogenesis of cacao: the effect of explant types and plant growth regulators. Indones J Agric Sci. 2019;20(2):69-76. doi:http//dx.doi.org/10.21082/ijas.v.20.n2.2019.p69–76
19. Daouda K, Modeste K, Oulo N, Edmond K. Induction of somatic embryos of recalcitrant genotypes of Theobroma cacao L. J Appl Biosci. 2019;133(1):13552. doi:10.4314/jab.v133i1.7
20. Eliane M, Modeste K, André S, Edmond K, Mongomaké K. Effect of Water Stress Induced by Polyethylene Glycol 6000 on Somatic Embryogenesis in Cocoa (Theobroma cacao L.). Agric Sci. 2019;10(09):1240-1254. doi:10.4236/as.2019.109092
21. Iracheta L, Cruz L, Lopez P, Avendaño C, Ortiz S. 2iP and brasinosteroids promote somatic embryogenesis induction in Theobroma cacao L. Agroproductividad. 2019;12(1):65-70. doi:https://doi.org/1010.32854/agrop.v0i0.1340
22. Maximova S, Alemanno L, Young A, Ferriere N, Traore A, Guiltinan M. Efficiency, genotypic variability, and cellular origin of primary and secondary somatic embryogenesis of Theobroma cacao L. Vitr Cell Dev Biol - Plant. 2002;38(3):252-259. doi:10.1079/IVP2001257
23. Fontanel A, Gire S, Labbe G, et al. In vitro multiplication and plant regeneration of Theobroma cacao L. via stable embryogenic calli. IAPTC Congr Plant Biotechnol 2002 beyond. Published online 2002:23-28.
24. Garcia C, Marelli J, Motamayor J, Villela C. Somatic Embryogenesis in Theobroma cacao L. In: Loyola-Vargas V, Ochoa-Alejo N, eds. Plant Cell Culture Protocols, Methods in Molecular Biology. Vol 1815. Springer Science; 2018:227-245. doi:https://doi.org/10.1007/978-1-4939-8594-4_15
25. Tapi A, Gnion M, Adiko A, Mahamadou S, Mboup C. Field Performance of Cocoa Somaclones Derived from Somatic Embryogenesis. J Plant Sci Agric Res. 2020;4(2):34. doi:10.36648/plant-sciences.4.1
26. Goenaga R, Guiltinan M, Maximova S, Seguine E, Irizarry H. Yield performance and bean quality traits of cacao propagated by grafting and somatic embryo-derived cuttings. HortScience. 2015;50(3):358-362. doi:10.21273/hortsci.50.3.358
27. Entuni G, Edward R, Nori H, Ahmad AK. Field performance of selected Malaysian cocoa clones regenerated through somatic embryogenesis cultures. Malaysian Appl Biol. 2018;47(1):97-102.
28. Guillou C, Fillodeau A, Brulard E, et al. Nestlé Cocoa plan: Cocoa propagation by somatic embryogenesis. In: Park YS, Bonga JM, eds. Proceedings of the Third International Conference of the IUFRO on “Woody Plant Production Integrating Genetic and Vegetative Propagation Technologies“. Vol 1. IUFRO; 2015:75-80.
29. Osorio T, Henao A, de la Hoz T, Urrea A. Propagation of IMC67 Plants, Universal Cacao (Theobroma cacao L.) Rootstock via Somatic Embryogenesis. Int J Fruit Sci. 2022;22(1):78-94. doi:10.1080/15538362.2021.2023067
30. Henao A, De-La-Hoz T, Ospina T, Garcés L, Urrea A. Evaluation of the potential of regeneration of different Colombian and commercial genotypes of cocoa (Theobroma cacao L.) via somatic embryogenesis. Sci Hortic (Amsterdam). 2018;229:148–156. doi:10.1016/j.scienta.2017.10.040
31. Urrea I, Atehortúa L, Gallego A. Regeneration through somatic embryogenesis of an elite colombian Theobroma cacao L. variety. Rev Colomb Biotecnol. 2011;XIII(2):39-50.
32. Montalbán I, García-Mendiguren O, Goicoa T, Ugarte M, Moncaleán P. Storage of plant material at low non-freezing temperature improves somatic embryogenesis in Pinus radiata. In: Proceedings of the Third International Conference of the IUFRO on “Woody Plant Production Integrating Genetic and Vegetative Propagation Technologies“. ; 2015:29-37.
33. Lelu-Walter MA, Thompson D, Harvengt L, Sanchez L, Toribio M, Pâques LE. Somatic embryogenesis in forestry with a focus on Europe: State-of-the-art, benefits, challenges and future direction. Tree Genet Genomes. 2013;9(4):883-899. doi:10.1007/s11295-013-0620-1
34. Barra-Jiménez A, Ruiz-Galea M, Celestino C, Alegre J, Toribio M. An overview of the current achievements in Holm oak (Quercus ilex L.) somatic embryogenesis. In: Park Y., Bonga JM, eds. Proceedings of the Third International Conference of the IUFRO on “Woody Plant Production Integrating Genetic and Vegetative Propagation Technologies“. IUFRO; 2015:105-107.
35. Savangikar V. Role of low cost options in tissue culture. In: Low Cost Options for Tissue Culture Technology in Developing Countries. FAO/IAEA Division of Nuclear Techniques in Food and Agriculture; 2004:11-15.
36. Ahloowalia B, Savangikar V. Low cost options for energy and labour. In: Low Cost Options for Tissue Culture Technology in Developing Countries. FAO/IAEA Division of Nuclear Techniques in Food and Agriculture; 2004:41-45.
37. Bhojwani SS, Dantu PK. Micropropagation. In: Bhojwani SS, Dantu PK, eds. Plant Tissue Culture: An Introductory Text. ; 2013:245-274. doi:10.1007/978-81-322-1026-9
38. Egertsdotter U, Ahmad I, Clapham D. Automation and scale up of somatic embryogenesis for commercial plant production, with emphasis on conifers. Front Plant Sci. 2019;10(February):1-13. doi:10.3389/fpls.2019.00109
39. Ahloowalia B, Prakash J, Savangikar V. Plant Tissue Culture. In: Low Cost Options for Tissue Culture Technology in Developing Countries. FAO/IAEA Division of Nuclear Techniques in Food and Agriculture; 2004:3-10.
40. George P, Jerold M. Low cost tissue culture technology for the regeneration of some economically important plants for developing countries Low Cost Tissue Culture Technology for the Regeneration of Some Economically Important Plants for Developing Countries. Int J Agric Environ Biotechnol Cit. 2013;6(December 2013).
41. Datta SK, Chakraborty D, Janakiram T. Low Cost Tissue Culture : An Overview. J Plant Sci Res. 2017;33(2):181-199. http://libaccess.mcmaster.ca/login?url=http://search.ebscohost.com/login.aspx?direct=true&db=eih&AN=126464537&site=ehost-live&scope=site
42. Etienne H, Breton D, Breitler JC, et al. Coffee somatic embryogenesis: How did research, experience gained and innovations promote the commercial propagation of elite clones from the two cultivated species? Front Plant Sci. 2018;871(November):1-21. doi:10.3389/fpls.2018.01630
43. Garcia C, Furtado de Almeida A, Costa M, et al. Abnormalities in somatic embryogenesis caused by 2,4-D: an overview. Plant Cell Tissue Organ Cult. 2019;137(2):193-212. doi:10.1007/s11240-019-01569-8
44. Das A, Mukherjee P, Jha TB. High frequency micropropagation of aloe vera L. Burm. f. as a low cost option towards commercialization. Plant Tissue Cult Biotechnol. 2010;20(1):29-35. doi:10.3329/ptcb.v20i1.5962
45. Pozoga M, Olewnicki D, Jabłońska L. In vitro propagation protocols and variable cost comparison in commercial production for Paulownia tomentosa × Paulownia fortunei hybrid as a renewable energy source. Appl Sci. 2019;9(11). doi:10.3390/app9112272
46. Sorensson C. Varietal pines boom in the US South. New Zeal J For. 2006;51(2):34-40.
47. Becerra E. Minimum wage in Colombia is one of the lowest compared to others in the Latin American region. La República. Published 2019. Accessed July 28, 2020. https://www.larepublica.co/alta-gerencia/salario-minimo-en-colombia-es-de-los-mas-bajos-en-comparacion-con-otros-de-la-region-2862119
48. Janekova J, Fabianova J, Fabian M. Assessment of economic efficiency and risk of the project using simulation. Int J Simul Model. 2019;18(2).
49. La Scalia G, Micale R, Miglietta PP, Toma P. Reducing waste and ecological impacts through a sustainable and efficient management of perishable food based on the Monte Carlo simulation. Ecol Indic. 2019;97:363-371.
50. Gallego A, Henao A, Urrea A, Atehortúa L. Polyphenols distribution and reserve substances analysis in cocoa somatic embryogenesis. Acta Biol Colomb. 2016;21(2):335-345. doi:http://dx.doi.org/10.15446/abc.v21n2.50196
51. Henao A, Salazar H, Urrea A. Quality of cocoa (Theobroma cacao L.) DNA from foliar tissue at different stages of development. Acta Agron. 2018;67(2):1-10. doi:10.15446/acag.v67n2.63046
52. Tomar UK, Negi U, Sinha AK, Danatu PK. An overview of the economic factors influencing micropropagation. In: Parihar P, Parihar L, eds. Advances in Applied Biotechnology. Agrobios. ; 2010:1-12.
53. Dong Y, Chen M, Wang X, Niu L, Fu Q, Xu Z. Establishment of in vitro regeneration system of woody oil crop Plukenetia volubilis. Mol Plant Breed. 2016;14(2):462-470.
54. Palisade. @RISK 8. Palisade Help Resources. Published 2020. Accessed October 16, 2020. https://help.palisade.com/v8/en/@RISK/@RISK.htm
55. Hoagland DR, Snyder WC. Nutrition of strawberry plant under controlled conditions:(a) effects of deficiencies of boron and certain other elements:(b) susceptibility to injury from sodium salts. In: Proc. Amer Soc. Hort. Sci. Vol 30. ; 1933:288941.
56. Zea R. Minagricultura announces renovation plan of 10,000 hectares of cocoa in 2021 to improve productivity. Ministerio de Agricultura y Desarrollo Rural. Published 2020. Accessed January 3, 2022. https://www.minagricultura.gov.co/noticias/Paginas/Minagricultura-anuncia-plan-de-renovación-de-diez-mil-hectáreas-de-cacao-en-2021-para-mejorar-la-productividad.aspx
57. Guillou C, Fillodeau A, Brulard E, et al. Indirect somatic embryogenesis of Theobroma cacao L. in liquid medium and improvement of embryo-to-plantlet conversion rate. Vitr Cell Dev Biol - Plant. 2018;54(4):377-391. doi:10.1007/s11627-018-9909-y
58. Norasekin T, Siti MA, Kasran R. Induction and propagation of somatic embryos from cell suspension cultures of Theobroma cacao L. Malaysian Cocoa J. 2020;12(November).
59. Natarajan N, Sundararajan S, Ramalingam S, Chellakan PS. Efficient and rapid in-vitro plantlet regeneration via somatic embryogenesis in ornamental bananas (Musa spp.). Biologia (Bratisl). 2020;75(2):317-326. doi:10.2478/s11756-019-00358-0
60. Kong EYY, Biddle J, Foale M, Adkins SW. Cell suspension culture: A potential in vitro culture method for clonal propagation of coconut plantlets via somatic embryogenesis. Ind Crops Prod. 2020;147(January):112125. doi:10.1016/j.indcrop.2020.112125
61. Awada R, Verdier D, Froger S, et al. An innovative automated active compound screening system allows high-throughput optimization of somatic embryogenesis in Coffea arabica. Sci Rep. 2020;10(1). doi:10.1038/s41598-020-57800-6
62. Bonga JM. A comparative evaluation of the application of somatic embryogenesis, rooting of cuttings, and organogenesis of conifers. Can J For Res. 2015;45(4):379-383. doi:10.1139/cjfr-2014-0360
63. Cervelli R, Senaratna T. Economic aspects of somatic embryogenesis. In: Aitken J, Kozai T, Lila M, eds. Automation and Environmental Control in Plant Tissue Culture. Springer Science; 1995:29-64. doi:10.1007/978-94-015-8461-6_3
64. Chu I. Economic analysis of automated micropropagation. In: Aitken J, Kozai T, Lila M, eds. Automation and Environmental Control in Plant Tissue Culture. Springer Science; 1995:19-27. doi:10.1007/978-94-015-8461-6_2
65. Suárez-Castellá M, Triana-Gutiérrez R, Pérez Z, Rodríguez M. Procedimiento metodológico para el cálculo de normas de trabajo de los operadores de cabinas de flujo laminar en la propagación masiva de plantas in vitro. Biotecnol Veg. 2006;6(4):195-202.
66. Chen C. Cost analysis of plant micropropagation of Phalaenopsis. Plant Cell Tissue Organ Cult. 2016;126(1):167-175. doi:10.1007/s11240-016-0987-4
67. Kaur A, Sandhu JS. High throughput in vitro micropropagation of sugarcane (Saccharum officinarum L.) from spindle leaf roll segments: Cost analysis for agri-business industry. Plant Cell Tissue Organ Cult. 2015;120(1):339-350. doi:10.1007/s11240-014-0610-5
68. Sahu J, Sahu RK. A Review on Low Cost Methods for <I>In vitro</I> Micropropagation of Plant Through Tissue Culture Technique. UK J Pharm Biosci. 2013;1(1):38. doi:10.20510/ukjpb/1/i1/91115
69. Sudipta K., Kumara M, Balasubramanya S, Anuradha M. Cost Efective Approach for in vitro Propagation of ( Leptadenia Reticulata Wight & ARN.) - A threatened Plant of Medicinal Importance. J Phytol. 2011;3(2):72-79.
70. Demo P, Kuria P, Nyende AB, Kahangi EM. Table sugar as an alternative low cost medium component for in vitro micro-propagation of potato (Solanum tuberosum L.). African J Biotechnol. 2008;7(15):2578-2584. doi:10.5897/AJB08.263
71. Prakash S, Hoque M., Brinks T. Culture media and containers. In: Low Cost Options for Tissue Culture Technology in Developing Countries. FAO/IAEA Division of Nuclear Techniques in Food and Agriculture; 2004:29-40.
72. von Aderkas P, Kong L, Prior N. In vitro techniques for conifer embryogenesis. Veg Propag For Trees Seoul, Korea Korea For Res Inst. Published online 2016:335-350.
73. Tikkinen M, Varis S, Peltola H, Aronen T. Improved germination conditions for Norway spruce somatic cotyledonary embryos increased survival and height growth of emblings. Trees - Struct Funct. 2018;32(6):1489-1504. doi:10.1007/s00468-018-1728-6
74. Llebrés MT, Pascual MB, Debille S, et al. The role of arginine metabolic pathway during embryogenesis and germination in maritime pine (Pinus pinaster Ait.). Tree Physiol. 2017;38(3):471-484. doi:10.1093/treephys/tpx133
75. Hazubska-Przybył T, Wawrzyniak M, Obarska A, Bojarczuk K. Effect of partial drying and desiccation on somatic seedling quality in Norway and Serbian spruce. Acta Physiol Plant. 2015;37(1):1-9. doi:10.1007/s11738-014-1735-1
76. Valverde C, Ramírez C, Kloepper JW, Cassán F. Current Research on Plant-Growth Promoting Rhizobacteria in Latin America: Meeting Report from the 2nd Latin American PGPR Workshop. J Plant Growth Regul. 2015;34(1):215-219. doi:10.1007/s00344-014-9470-9
77. Pila L, Freitas H, do Nascimento L, Guerra M. Epigenetics of long-term somatic embryogenesis in Theobroma cacao L.: DNA methylation and recovery of embryogenic potential. Plant Cell Tissue Organ Cult. 2017;131(2):295-305. doi:10.1007/s11240-017-1284-6
78. BIOBAHIA. Biofábrica de Bahía - Plantas de cacao. Plantas de cacao. Published 2020. Accessed November 26, 2020. https://www.biofabrica.org.br/nossa-historia
Received: 14 July 2021 / Accepted: 10 December 2021 / Published:15 May 2022
Citation: Henao-Ramírez, AM.; Palacio- Hajduk, DH.; Urrea-Trujillo, AI. Cost Analysis of Cacao (Theobroma cacao L.) Plant Propagation through the Somatic Embryogenesis Method. Revis Bionatura 2022;7(2) 2. http://dx.doi.org/10.21931/RB/2022.07.02.2