2018.03.01.9
Files > Volume 3 > Vol 3 No 1 2018
INVESTIGATION / RESEARCH
Evaluación de los Hongos Micorrízicos Arbusculares de zonas del trópico húmedo del Ecuador
Evaluation of Arbuscular Mycorrhizal Fungi from humid tropical areas of Ecuador

Emy Moína-Quimí, Rodrigo Oviedo-Anchundia, Shayler Nieto-Barciona, Paúl Herrera-Samaniego, Milton Barcos-Arias.
Available from: http://dx.doi.org/10.21931/RB/2018.03.01.9
RESUMEN
El estudio se enfocó en el análisis de la simbiosis entre hongos micorrízicos arbusculares y árboles del trópico húmedo ecuatoriano, mediante técnicas físicas de separación se aisló esporas de hongos micorrízicos arbusculares para obtener la densidad por gramos de suelo, mientras que el porcentaje de micorrización en la flora representativa fue determinada al comprobar la presencia de estructuras micorrízicas en el interior de las raíces de las plantas evaluadas, y además, se realizó un análisis físico-químico del suelo estudiado. Los análisis arrojaron resultados que demuestran una densidad media de esporas, también se determinó un porcentaje de micorrización por arriba del 50% en las floras representativas. A nivel de esporas, se identificaron géneros predominantes de las zonas, estando presentes Acaulospora, Ambispora, Claroideoglomus, Diversispora, Entrophospora, Funneliformis,Gigaspora, Glomus, Pacispora y Scutellospora, de los cuales se llegó a identificar por métodos taxonómicos cerca de 15 especies. Concluyendo que existe una diversidad de hongos micorrícicos arbusculares en el trópico húmedo ecuatoriano.
Palabras-claves: Simbiosis, micorrización, condiciones abióticas, hospedador, hongos, trópico húmedo ecuatoriano.
ABSTRACT
The study approached the symbiotic analysis among Arbuscular Mycorrhizal Fungi and trees of Ecuadorian humid tropics, through physical separation techniques spores were isloted to determinate the Arbuscular Mycorrhizal Fungi density per soils gram, also the mycorrhization percentage of representative flora was determinated by the presence of Mycorrhizal structures inside the evaluated plants roots, additionally, it was performanced a physical-chemistry analysis of soil. The different analyzes showed an average density of spores, a mycorrhization percentage over 50 % in the representative flora. The predominant genera of spores were taxonomically identified as Acaulospora, Ambispora, Claroideoglomus, Diversispora, Entrophospora, Funneliformis, Gigaspora, Glomus,Pacispora and Scutellospora, there has been about 15 species in selected zones. Concluding that there is a great diversity of arbuscular mycorrhizal fungi in the ecuadorian humid tropics.
Key-words: Symbiosis, micorrization, abiotic conditions, host, fungi, ecuadorian humid tropics.
INTRODUCCIÓN
La rizósfera es el ambiente donde ocurren las principales interacciones entre los microorganismos y las plantas. Gran parte de estas interacciones afectan al desarrollo, productividad y funciones esenciales de la flora circundante. Además, en este nivel se produce la descomposición de la materia orgánica con la colaboración de los microorganismos presentes en la rizósfera, que a su vez desencadena en la liberación de los nutrientes al suelo1. Otros microorganismos rizosféricos se encargan de la captación de los nutrientes liberados, es en este grupo donde encontramos a los hongos micorrízicos arbusculares (HMA).
Los HMA son simbiontes obligados que colonizan, o micorrizan, aproximadamente el 74% de plantas terrestres2,rasgo característico que les permite adaptarse en diferentes ambientes, llegando a comprender del 5 al 50% de la biomasa microbiana total del suelo3. La relación simbiótica entre planta y hongo les otorga grandes beneficios a ambas partes involucradas. Las plantas generan las principales fuentes del carbono necesario para el desarrollo y la multiplicación de los HMA. Por otra parte, los HMA confieren a las plantas hospedadoras una mejor absorción y toma de nutrientes, así como la captación de iones inmóviles de fósforo. Adicionalmente, la interacción con los HMA les otorga mayor resistencia a los diferentes tipos de estrés biótico y abiótico, y aumenta la productividad y el desarrollo radicular de la planta gracias a la formación de microagregados del suelo4.
Si bien los HMA no tienen especificidad por colonizar una especie de planta en particular, esto no significa que haya un grado de compatibilidad idéntico entre todas las interacciones de especies de HMA y las plantas terrestres. Se ha comprobado que la compatibilidad de la interacción simbiótica entre plantas y HMA es mayor entre unos géneros que entre otros, y esta peculiaridad puede ser aprovechada en muchos ámbitos5. Los países desarrollados han creado tecnologías y productos a base de micorrizas, sin embargo, esta interacción no ha sido estudiada en su totalidad.
En Ecuador, el desarrollo biotecnológico utilizando HMA ha ido acrecentándose, sin embargo, cabe mencionar la poca información existente respecto a la identificación de los principales géneros de HMA presentes en los bosques ecuatorianos, siendo esta información necesaria para la aplicación y elaboración de productos a base de HMA. Así mismo sería importante conocer el número más probable de propágulos micorrizales infectivos6. Respecto a esta problemática, este estudio quiere dar a conocer los géneros predominantes de estos hongos y las condiciones a las cuales están expuestos naturalmente en bosques del trópico húmedo, teniendo en cuenta que son uno de los sectores que posee mayor diversidad biológica en el país7.
MATERIALES Y MÉTODOS
La evaluación de los HMA fue realizada en los cantones Pallatanga, Bucay y La Maná, estos tres sitios poseen una vegetación y condiciones climáticas similares, presentando variabilidad en sus climas desde cálido, templado y hasta frio, con precipitaciones variable entre 1000-3000 mm según la época del año, lo cual es propio del trópico húmedo ecuatoriano8,9. En estos cantones se seleccionaron sectores específicos para hacer muestreos en la temporada de invierno, recolectando muestras de suelo rizosférico y raíces de plantas principalmente de banano (Musa spp.), cacao (Theobroma cacao), y de especies forestales como el niguito (Muntingia calabura). Las coordenadas de referencia tomadas en los sectores A1- A2 de Pallatanga fueron 02°09'26.0'' S y 079°03'45.2'' W,en los sectores B1 – B2 en Bucay fueron 02°11'59.2'' S y 079°07'06.8'' W, y en los sectores C1 – C2 en La Maná fueron 00°56'32.0'' S y 079°14'16.5'' W. El suelo rizosférico y las raíces fueron aprovechados para determinar el porcentaje de micorrización, la densidad de esporas por gramos de suelo, e identificar los principales géneros de esporas HMA de la zona. Además, con estas muestras se determinaron el análisis físico-químico de los suelos.
Análisis físico-químico de los suelos
El análisis físico-químico de las muestras fue realizado en el departamento de suelo del Instituto Nacional de Investigaciones Agropecuarias (INIAP). Por cada muestra se envió 1 Kg de suelo correspondiente a cada sector seleccionado, y se determinó la textura y la humedad del suelo, la materia orgánica por el método de Walkley-Black, fósforo total, amonio y potasio mediante Olsen modificado pH 8.5, el pH en suspensión suelo: agua 1:2.5, y la conductividad eléctrica en pasta saturada.
Aislamiento, determinación de densidad e identificación de los hongos micorrízicos arbusculares
La técnica empleada para el aislamiento de esporas, corresponde a la metodología de tamizado en húmedo y decantación10, continuando con la extracción por gradientes de densidad11. El sobrenadante final fue disuelto en agua para lavar parcialmente a las esporas, y el criterio para determinar la densidad de esporas HMA fue el siguiente criterio: densidad baja corresponde < 1 esporas/g de suelo; densidad media corresponde 1 – 10 esporas/g de suelo; y densidad alta corresponde > 10 esporas/g de suelo12.
Posterior al proceso de aislamiento de esporas, éstas fueron fijadas en placas portaobjetos para su conservación y posterior identificación. La técnica de fijación consistía en el uso de PVLG (Alcohol polivinílico-ácido láctico-glicerol) y reactivo de Melzer13. Además, se tomó en cuenta durante el montaje de esporas, que el uso de PVLG + reactivo de Melzer sería empleado para esporas en mal estados, mientras que el PVLG sería aplicado para esporas en buen estado. Aproximadamente, 15 esporas fueron colocadas por placa.
Determinación del porcentaje de micorrización en raíces
Se tomaron 10 individuos de las especies agroforestales predominantes como banano, cacao y niguito por cada sector, de estos fueron extraídas las raíces que se encontraban a una profundidad de 5 - 20 cm. El análisis consistió en comprobar la presencia de estructuras micorrízicas (hifas, vesículas y arbúsculos) dentro de las raíces. Las muestras pasaron por los procesos de clarificación y tinción14, luego se colocó 10 raíces de 1,5 cm de longitud en placas porta objetos.
Las raíces fueron analizadas bajo el lente 20X de un microscopio óptico compuesto, dividiendo a las raíces en tres campos ópticos observables, y a cada campo se lo diagnosticó como positivo si se observaba al menos una de las tres estructuras micorrízicas. La fórmula utilizada para determinar el porcentaje aproximado de micorrización fue:
Identificación morfológica de las esporas de hongos micorrízicos arbusculares
La identificación de los géneros predominantes de HMA se basó en las características morfológicas de las esporas, principalmente en el color, la textura, el tamaño, el número de paredes y capas que recubren a la espora, la cantidad de cicatrices presentes, y la unión de la hifa suspensoria a la espora. Las características fueron examinadas empleando manuales morfológicos de libre acceso, dos de ellos son manuales en línea, uno propuesto por INVAM de la Universidad de Virginia16, otro manual de libre acceso fue propuesto en la página web de Blaszkowski17, y el tercer manual corresponde al anexo de una tesis doctoral que empleó la recopilación de claves taxonómicas y artículos científicos para la determinación de especies HMA18.
RESULTADOS Y DISCUSIÓN.
Caracterización físico-químico del suelo
De acuerdo a los resultados expuestos en la tabla 1, el tipo de textura que presentan los suelos colectados es franco-arenoso, el pH varía según el sector encontrándose desde 6,8 hasta 5, que corresponden a pH neutro y fuertemente ácido respectivamente, según la norma NOM-021-RECNAT-200019, siendo a su vez el factor pH de gran relevancia para la adaptación de los HMA, prefiriendo pH neutros, aunque pueden llegar a desarrollarse hasta en pH ácidos20. Mientras que la conductividad eléctrica los valores oscilaron entre 0.23 y 1,10 mS/cm que corresponden a niveles bajo en salinidad, estas concentraciones no afectan a la micorrización ya que está demostrado que en suelos salinos puede progresar la simbiosis21,22. En lo que respecta a los niveles de P también variaron según el sector encontrándose entre 11,0 a 30,0 µg/ml considerandos como nivel medio según reporte del análisis. La simbiosis entre HMA y las plantas se inhibe cuando los niveles de P son altos5, los niveles bajos y medios de P favorecen la relación simbiótica de HMA y planta23.
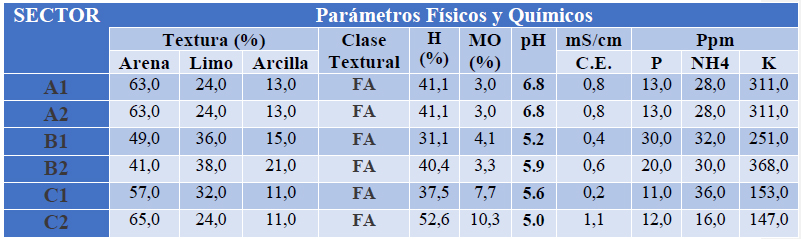
Tabla 1.- Análisis físico-químico de las diferentes muestras de suelo colectadas en sectores del trópico húmedo. “FA” hace referencia a la textura FRANCO-ARENOSA del suelo.
Otros parámetros tomados en cuenta fueron el nivel de nitrógeno (N), la humedad del suelo y el porcentaje de materia orgánica (MO), con el fin de conocer si éstos influyen en la micorrización de las plantas en los sectores seleccionados. Estudios realizados por Martínez y Pugnaire24 mencionan que la relación entre N y HMA tiene mayor incidencia en la micorrización que la relación de ésta frente al P, esta afirmación se apoya por estudios que han comprobado que el nivel de N es inversamente proporcional a la micorrización en plantas, mientras menor sean los niveles de N, mayor sería micorrización por HMA25,26. En base a la información citada, los niveles de N reportados en la tabla 1 no afectarían a la simbiosis de HMA en las zonas analizadas. Respecto a la humedad y el porcentaje de MO, la humedad en el suelo no sobrepaso el 52%, y en MO varió entre 3% y 10%. También, ha sido reportado que la adaptación de los HMA es más favorable en suelos húmedo, que en suelos inundado o secos27, sin embargo otros estudios realizados en bosques del trópico húmedo, muestran que la humedad del suelo como factor ambiental, influye muy poco en la frecuencia de la micorrización28. Con relación al porcentaje de MO, un estudio realizado en el caribe colombiano, la MO y su descomposición afecta directamente a los niveles de pH en el suelo, y como ya fue mencionado el pH tiene un efecto en la micorrización29.
Cuantificación de la densidad de esporas HMA en suelo rizosférico.
La densidad de esporas HMA nativas puede estar influenciada principalmente por aspectos ambientales18,30. Los resultados exhibidos en la tabla 2, muestran la cantidad de esporas por gramos de suelo correspondiente a los diferentes sectores seleccionados. Los resultados revelan un promedio de densidad media que oscila entre 1,4 esporas/gramo de suelo y 3,4 esporas/gramo de suelo. Las muestras que presentan mayor densidad correspondes al sector C2 ubicado en el cantón La Maná, y las muestras con menor densidad corresponden al sector B2 ubicado en el cantón Bucay. En todos los sectores estudiados, la densidad de esporas por gramos de suelo es media. Se prevé que la tenue diferencia de densidad de esporas exhibidas entre los cantones, es dada por los distintos niveles de pH, humedad, materia orgánica y nutrientes a los que están expuestos.
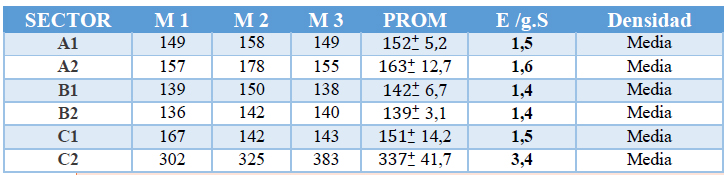
Tabla 2.- Densidad de esporas en suelo rizosférico. Donde (M) corresponden al número de réplica y (PROM), al promedio de las tres réplicas, y E/G.S es la relación de esporas por g de suelo.
Determinación del porcentaje de colonización en las raíces de la flora representativa.
Los resultados descritos en la figura 1, corresponde al análisis efectuado a raíces de la flora representativa en cada sector de estudio. Al colocar 10 raíces por placa, dio como resultado un mínimo de 30 campos observables, en ellos figuraron las estructuras hifas, vesículas y arbúsculos. En la figura 2 están representadas las estructuras propias de los HMA y en la figura 1 queda expuesto, que los sectores con mayor porcentaje de colonización en sus raíces, corresponden a A1 y C1, ubicado en los cantones Pallatanga y La Maná respectivamente. Los valores mínimos de colonización corresponden a los sectores A2 y B2, lugares que han estado expuestos en baja medida, a intervenciones antropogénicas por parte de los habitantes del sector. Aun así, se considera a la flora representativa como micorrizada, debido a que el porcentaje de colonización está por encima del 50%.
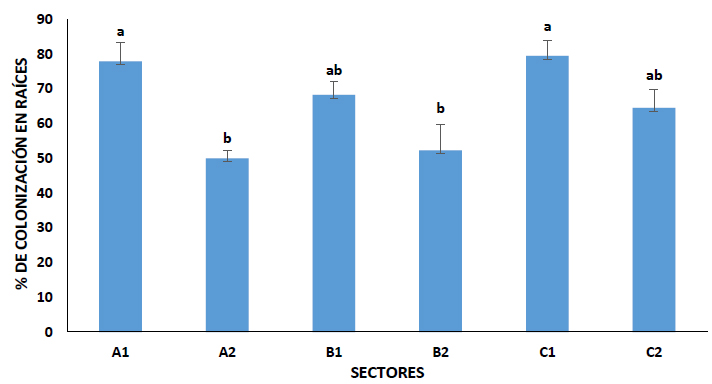
Figura 1.- Porcentaje de micorrización en raíces de la flora representativa correspondiente a los sectores seleccionados. Diferentes letras en las columnas indican diferencias estadísticas significativas (Tukey, α=0.05) n=8.
Bago31, menciona la diferencia entre efectividad e infectividad en una planta, mostrando que una planta con un alto nivel de porcentaje micorrización puede tener efectos poco notables, mientras que la misma planta micorrizada por otras especies de HMA aún con niveles más bajos de micorrización, pueden presentar mayor efecto positivo en la misma. Por otra parte, el porcentaje de micorrización o colonización en las raíces muestreadas no está relacionado específicamente con la densidad de esporas HMA, este comportamiento también fue observado en un estudio dado en la Amazonía Colombiana32, donde el comportamiento de micorrización podría ser mayor en una especie que en otra, sin embargo el número de esporas podría ser no muy alto en dicho tratamiento.
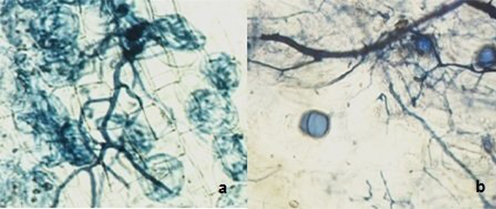
Figura 2.- Micorrización en raíces. a) Colonización por hifas y arbúsculos; b) Colonización por hifas y vesículas.
Identificación de los principales géneros de HMA en las zonas del trópico húmedo.
Algunos géneros de HMA fueron hallados en los lugares evaluados en el trópico húmedo ecuatoriano, alrededor de 15 especies fueron encontradas en los principales géneros como se muestra en la figura 3. Los géneros encontrados en mayor abundancia corresponden a Acaulospora, Ambispora, Claroideoglomus, Diversispora, Funneliformis,Gigaspora, Glomus, Pacispora, y Scutellospora. Varias especies de HMA que antes fueron definida comoEntrophospora, han sido ubicadas recientemente en el género Acaulospora, dejando solo una especie enEntrophospora16, denominada Entrophospora infrequens33 y ésta fue hallada en el presente estudio.
El género Acaulospora fue identificado debido a las cicatrices presentadas en la superficie de la espora, característica propia de su género al desprenderse la espora de la hifa suspensoria, además, en su gran mayoría poseen una superficie ornamentada, aunque hay especies las cuales poseen una superficie lisa18. Las Ambisporasposeen como característica principal, el desarrollo de dimórfico de esporas glomoides y acaulosporoides34, se presume que las especies de Ambispora encontradas son acaulosporoides, ya que presentan características muy similares con las Acaulosporas. Del género Claroideoglomus, se logró identificar una de las especies de HMA debido a la inminente presencia de una capa evanescente en el exterior de la espora, y en el interior una pared semiflexible35.

Figura 3.- Esporas HMA representativas de los sectores seleccionados. a) Acaulospora alpina; b) Acaulospora scrobiculata; c) Acaulospora spinosa; d) Entrophospora infrequens; e) Diversispora tortuosa; f) Diversispora celata; g) Diversispora sp.; h) Pacispora robiginia; i) Gigaspora margarita; j) Scutellospora sp.; k) Ambispora sp. [1]; l) Ambispora sp. [2]; m) Funneliformis geosporus; n) Claroideoglomus lamellosum; o) Glomus sp. [1]; p) Glomus sp. [2].
Las esporas de los géneros Diversispora, Funneliformis, Glomus y Pacispora tienen un origen de germinación muy parecido, sin embargo, difieren en tamaño, agrupación, color y número de capas18, factores que en este caso hicieron posible su identificación y clasificación. Tanto los géneros Gigaspora y Scutellospora fueron reconocidos por el tamaño de sus esporas y por el origen a partir de células bulbosas, ambos géneros presentan esporas de gran tamaño, pero poseen característica que difieren entre ellas, un ejemplo claro es que las Gigasporas halladas poseen una pared externa totalmente lisa18, mientras que en el género Scutellospora está reportado la existencia de esporas con pared lisas y ornamentadas, además las Scutellospora al ser aplastada durante el proceso de preservación, demostraron 3 paredes germinales en el interior de la espora, propias de este género36.
Especies de los géneros Acaulospora, Glomus, Gigaspora y Scutellospora, ya han sido reportadas anteriormente en sistemas agroforestales del Ecuador28, y de las esporas correspondientes a estos géneros, se llegó a identificar ciertas especies de HMA, sin embargo, se presume que la especie Acaulospora alpina es reportada por primera vez en zonas del trópico ecuatoriano. Una especie aislada en este estudio fue identificada como Gigaspora margarita, se conoce poco de la presencia de esta especie en Ecuador, y del género Scutellospora no se logró identificar las especies halladas. Sin embargo hay mención del género Gigaspora y Scutellospora en estudios realizados a palmito en Santo Domingo de los Tsachilas37, y en otros estudios realizados en la provincia de los Ríos, que también demuestran la presencia de Gigaspora y Scutellospora en sus muestras de suelo38.
En Latinoamérica, la Diversispora tortuosa ha sido reportada en estudios realizados a monocultivos de maíz en Brasil39, y la Diversispora celata, junto a las especies Funneliformis geosporus, y Entrophospora infrequens han sido reportadas en un estudio doctoral realizado en Colombia40, al igual que especies de los génerosClaroideoglomus, Ambispora y Pacispora, que si bien no han sido identificados en su totalidad, se conoce de su presencia en los estudios ya mencionados.
CONCLUSIONES
Los niveles medio de fósforo y nitrógeno al igual que los valores de pH, materia orgánica y conductividad eléctrica presentes en las zonas de estudios son adecuados para la proliferación de HMA. Sin embargo, se encontró una baja densidad de esporulación en los suelos rizosféricos analizados. Mientras que el porcentaje de micorrización total en raíces estuvo por encima del 50% en todos los sitios estudiados. En lo que respecta a la diversidad se identificaron 10 géneros y 15 especies de HMA en los lugares evaluados, lo que pone de manifiesto la diversidad de estos microorganismos en el trópico húmedo ecuatoriano.
REFERENCIAS BIBLIOGRAFÍCAS
1. Van Der Heijden, M. G. A., Bardgett, R. D. & Van Straalen, N. M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 11, 296–310 (2008). doi: 10.1111/j.1461-0248.2007.01139.x
2. Wang, B. & Qiu, Y.-L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza16, 299–363 (2006). doi: 10.1007/s00572-005-0033-6
3. Vertel, M. & Pérez, A. Evaluación de la colonización de micorrizas arbusculares en pasto Bothriochloa pertusa ( L ) A . Camus. Rev. MVZ Córdova 15, 2165–2174 (2010). http://www.redalyc.org/articulo.oa?id=69318985004
4. Van der Heijden, M. et al. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Lett. to Nat. 396, 69–72 (1998). doi: 10.1038/23932
5. Barrer, S. E. El Uso de Micorrízicos Arbusculares Como Una Alternativa Para La Agricultura. Rev. Chil. Hist. Nat. 7, 124–126 (2009). http://www.scielo.org.co/pdf/bsaa/v7n1/v7n1a14.pdf
6. Porter, W. The ‘most probable number’ method for enumerating infective propagules of vesicular arbuscular mycorrhizal fungi in soil. Aust. J. Soil Res. 17, 515–519 (1979). doi: 10.1071/SR9790515
7. Bravo Velásquez, E. La biodiversidad en el Ecuador. (Universidad Politécnica Salesiana, 2014). http://dspace.ups.edu.ec/bitstream/123456789/6788/1/La Biodiversidad.pdf
8. Alarcón, P. Perfil Territorial del Cantón Pallatanga. (2013). http://repositorio.cedia.org.ec/bitstream/123456789/853/1/Perfil territorial PALLATANGA.pdf
9. Centro del Agua y Desarrollo Sustentable. Proyecto de Evaluación de Vulnerabilidad y Reducción de Riesgo de Desastres a Nivel Municipal en el Ecuador. (2013). https://www.google.com.ec/url?sa=t&rct=j&q=&esrc=s&source=web&cd=1
10. Gerdemann, J. W. & Nicolson, T. H. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 46, 235–244 (1963). doi: 10.1016/S0007-1536(63)80079-0
11. Furlan, V., Bartschi, H. & Fortin, J.-A. Media for density gradient extraction of endomycorrhizal spores.Trans. Br. Mycol. Soc. 75, 336–338 (1980). doi: 10.1016/S0007-1536(80)80102-1
12. Guerra S., B. E. & Chacón V., M. R. Simbiosis micorrízica arbuscular y acumulación de aluminio en Brachiaria decumbens y Manihot esculenta. Biotecnol. en el Sect. Agropecu. y Agroindustrialtrial 10, 87–98 (2012). http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S1692-35612012000200011&lng=en&nrm=iso
13. Koske, R. E. & Tessier, B. A Convenient, Permanent Slide Mounting Medium. Mycol. Soc. Am. Newsl. 34,59 (1983). http://ccb.ucr.edu/soillabfiles/labprotocol/slide_mounting.doc.
14. Phillips, J. M. & Hayman, D. S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 55, 158–161 (1970). doi: 10.1016/S0007-1536(70)80110-3
15. McGonigle, T. P., Miller, M. H., Evans, D. G., Fairchild, G. L. & Swan, J. a. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 115, 495–501 (1990). doi: 10.1111/j.1469-8137.1990.tb00476.x
16. Morton, J. B. International Culture Collection of (Vesicular) Arbuscular Mycorrhizal (INVAM) - West Virginia University. INVAM (2000). Available at: http://invam.wvu.edu/.
17. Blaszkowski, J. Arbuscular mycorrhizal fungi (Glomeromycota), Endogane and Complexipes species deposited in the Department of Plant Pathology, University of Agriculture in Szczecin, Poland. ZOR (2003). Available at: http://www.zor.zut.edu.pl/Glomeromycota/index.html.
18. Méndez Cortés, H. Diversidad de hongos micorrícicos arbusculares asociados al cedro rojo (Cedrela odorata L.) en dos ecosistemas tropicales de Veracruz, México. (UNIVERSIDAD AUTÓNOMA DE NUEVO LEÓN, 2012). http://eprints.uanl.mx/id/eprint/3102
19. SEMARNAT. Especificaciones de fertilidad, salinidad y clasificación de suelos, estudio, muestreo y análisis. Secretaria de medio ambiente y recursos naturales. Norma Oficial Mexicana NOM-021-RECNAT-2000. 15–16 (2002). http://biblioteca.semarnat.gob.mx/janium/Documentos/Ciga/libros2009/021.pdf
20. Garzón, L. Importancia de las micorrizas arbusculares (MA) para un uso sostenible del suelo en la amazonia colombiana. Luna Azul 42, 217–234 (2016). http://www.cazalac.org/mwar_lac/fileadmin/document
21. Hajiboland, R., Aliasgharzadeh, N., Farsad Laiegh, S. & Poschenrieder, C. Colonization with arbuscular mycorrhizal fungi improves salinity tolerance of tomato ( Solanum lycopersicum L .) plants. Plant Soil 331, 313–327 (2010). doi: 10.1007/s11104-009-0255-z
22. Heikham, E., Bhoopander, G. & Kapoor, R. Ultrastructural evidence for AMF mediated salt stress mitigation in Trigonella foenum-graecum. Mycorrhiza 23, 71–86 (2013). doi: 10.1007/s00572-012-0449-8
23. Habte, M. & Osorio, N. W. Arbuscular Mycorrhizas : Producing and Applying Arbuscular Mycorrhizal Inoculum. Coll. Trop. Agric. Hum. Resour. 47 (2001). ISBN: 192932510X. https://www.ctahr.hawaii.edu/oc/freepubs/pdf/SCM-5.pdf
24. Martínez, L. B. & Pugnaire, F. I. Interacciones entre las comunidades de hongos formadores de micorrizas arbusculares y de plantas. Algunos ejemplos en los ecosistemas semiáridos. Ecosistemas 18, 44–54 (2009). http://revistaecosistemas.net/index.php/ecosistemas/article/view/65
25. Santos, J. C., Finlay, R. D. & Tehler, A. Molecular analysis of arbuscular mycorrhizal fungi colonising a semi-natural grassland along a fertilisation gradient. New Phytol. 172, 159–168 (2006). doi: 10.1111/j.1469-8137.2006.01799.x
26. Fitzsimons, M. S., Miller, R. M. & Jastrow, J. D. Scale-dependent niche axes of arbuscular mycorrhizal fungi. Oecologia 158, 117–127 (2008). doi: 10.1007/s00442-008-1117-8
27. Miller, S. P. Arbuscular mycorrhizal colonization of semi-aquatic grasses along a wide hydrologic gradient.New Phytol. 145, 145–155 (2000). doi: 10.1046/j.1469-8137.2000.00566.x
28. Prieto-Benavides, O. O. et al. Identificación de hongos micorrízicos arbusculares en sistemas agroforestales con cacao en el trópico húmedo Ecuatoriano. Agron. Mesoam. 23, 233–239 (2012). http://www.redalyc.org/articulo.oa?id=43724664002
29. Perez, C. A., Rojas, S. J. & Montes, V. . D. Hongos formadores de micorrizas arbusculares: una alternativa biologica para la sostenibilidad de los agroecosistemas de praderas en el caribe colombiano. Rev Colomb. Cienc. Anim 3, 366–385 (2011). http://dialnet.unirioja.es/servlet/oaiart?codigo=3817504
30. Su, Y.-Y., Sun, X. & Guo, L.-D. Seasonality and host preference of Arbuscular Mycorrhizal Fungi of five plant species in the inner Mongoloia steppe, China. Brazilian J. Microbiol. 42, 57–65 (2011). doi: 10.1590/S1517-83822011000100008
31. Bago, B., Pfeffer, P. E. & Shachar-Hill, Y. Carbon metabolism and transport in arbuscular mycorrhizas.Plant Physiol. 124, 949–58 (2000). doi: 10.1104/pp.124.3.949
32. León Velandia, D. Evaluación y caracterización de micorrizas arbusculares asociadas a yuca (Manihot esculenta sp) en dos regiones de la Amazonía Colombiana. (Pontifica Universidad Javeriana, 2006). http://www.javeriana.edu.co/biblos/tesis/ciencias/tesis296.pdf
33. Sieverding, E. & Oehl, F. Revision of Entrophospora and description of Kuklospora and Intraspora, two new genera in the arbuscular mycorrhizal Glomeromycetes. J. Appl. Bot. Food Qual. 80, 69–81 (2006). http://pub.jki.bund.de/index.php/JABFQ/article/view/2192
34. Walker, C. et al. Molecular phylogeny and new taxa in the Archaeosporales (Glomerycota): Ambispora fennica gen. sp. nov., Ambisporaceae fam. nov., and emendation of Archaeospora and Archaeosporaceae. Mycol. Res. 111, 137–53 (2007). doi: 10.1016/j.mycres.2006.11.008
35. Schüßler, A. & Walker, C. The Glomeromycota. A species list with new families and new genera. The Royal Botanic Garden Edinburgh 22, 24–26 (2010). http://www.amf-phylogeny.com/species_infos/higher_taxa/funneliformis_claroideoglomus_rhizophagus_redeckera.pdf
36. Walker, C. & Sanders, F. E. Taxonomic concepts in the Endogonaceae : III . The separation of Scutellospora gen . nov . from Gigaspora Gerd. & Trappe. Mycotaxon 27, 169–182 (1986). https://www.researchgate.net/publication/285329919
37. Paillacho Cedeño, F. I. Evaluación de la efectividad de las micorrizas arbusculares nativas sobre el desarrollo y estado nutritivo del palmito (Bactris gasipaes HBK) en esta de vivero, en Santo Domingo de los Tsachilas. (Escuela Politécnica del Ejército, 2010). http://repositorio.espe.edu.ec/bitstream/21000/2892/1/T-ESPE-IASA II-002332.pdf
38. Prieto Benavides, O. et al. Inoculación de Brachiaria decumbens con hongos formadores de micorriza arbuscular nativos del trópico húmedo ecuatoriano. Rev. Cienc. y Tecnol. 4, 9–18 (2011). http://www.uteq.edu.ec/revistacyt/publico/archivos/C2_Inoculación Brachiaria.pdf
39. Salas Méndez, D. F. Diversidade de fungos micorrízicos arbusculares e sua relação com atributos do solo em área de milho sob monocultivo e em consórcio com forrageiras no cerrado. (Universidade de Brasilía, 2016). http://repositorio.unb.br/handle/10482/21425
40. Ramírez Gómez, M. M. Evaluación de la diversidad de Hongos Formadores de Micorrizas Arbusculares (HFMA) y su relación con el establecimiento de simbiosis con Physalis peruviana L. (Universidad Nacional de Colombia, 2014). http://www.bdigital.unal.edu.co/40192/1/41706338.2014.pdf
Recibido: 30 agosto 2017
Aprobado: 28 enero 2018
Aprobado: 28 enero 2018
Emy Moína-Quimí 1, Rodrigo Oviedo-Anchundia1, Shayler Nieto-Barciona2, Paúl Herrera-Samaniego1, Milton Barcos-Arias1
1 Escuela Superior Politécnica del litoral, ESPOL, Facultad de Ciencias de la Vida, FCV, Centro de Investigaciones Biotecnológicas del Ecuador, CIBE, Campus Gustavo Galindo Km. 30.5 Vía Perimetral, P.O. Box 09-01-5863, Guayaquil, Ecuador.
2 Fundación Privada Ecuatoriana, Ave. Fco. De Orellana Ma111 Edif. Wordl Trade Center Torres B Piso 3 oficina 319, Guayas, Ecuador.