2019.04.01.4
Files > Volume 4 > Vol 4 No 1 2019
INVESTIGATION / RESEARCH
AAV Purification by anion-exchange chromatography
Purificación de AAV mediante cromatografía de intercambio aniónico.

Camacho, F1., Cerro, R.P2., Varas, N1., Leiva, M.J2., Toledo, J.R2. and Sánchez. O1.
Available from: http://dx.doi.org/10.21931/RB/2019.04.01.4
ABSTRACT
We describe a new optimized, scalable and reproducible method based on anion exchange chromatography to obtain high titers of rAAV vectors without empty capsids contamination. The method takes advantage of Q-sepharose matriz to development a scalable procedure. After the virus harvest from supernatant and lysate cells, virus crude was subjected to anion exchange chromatography with Q-sepharose column. Three different protocols were tested, and the elution peaks were evaluated through qPCR rAAV titration and 260/280 nm ratio determination in order to identified empty capsid-containing fractions. A 150 mM NH4Ac wash step fallowing by 1 M NaCl elution step generate a a high titer eluted fraction of rAAV with 1.334 260/280 nm ratio. The described method makes rAAV vector purification an easily adapted for a large scale GMP production format to produce empty capsid free rAAV for clinics.
Keywords: rAAV purification, anio-exchange, empty capsid
RESUMEN
Este trabajo describe un nuevo método optimizado, escalable y reproducible basado en la cromatografía de intercambio aniónico para obtener títulos elevados de vectores rAAV sin contaminación de cápsides vacías. El método aprovecha la matriz Q-sepharose para desarrollar un procedimiento escalable. Después de la recolección del virus del sobrenadante de cultivo y del lisado, el crudo viral se sometió a cromatografía de intercambio aniónico con una columna de Q-sefarosa. Se probaron tres protocolos diferentes, y los picos de elución se evaluaron mediante qPCR rAAV y la determinación de la relación de 260/280 nm para identificar las fracciónes que contienen cápsides vacías. Un paso de lavado con NH4Ac 150 mM seguido de un paso de elución con NaCl 1 M genera una fracción eluida de alto título de rAAV con una relación 260/280 nm de 1.334. El método descrito hace que la purificación del vector de rAAV se adapte fácilmente a un formato de producción de GMP a gran escala para producir rAAV libre de cápside vacío para clínicas.
Palabras claves: Purificación de rAAV, intercambio aniónico, capsidas vacias
INTRODUCTION
Recombinant Adeno-associated virus (rAAV) are simple virus with linear single-stranded DNA genome. rAAV have proved significant potential as DNA-delivery vectors. Their main advantages include a wide tropism, poor immunogenicity and the ability to maintain the transgene expression for long periods 1.
Since the introduction of Baculovirus expression vectors to produced high titers of rAAV, the problem of scalable production seems to be solved 2. However, manufacture methods that enable to obtain rAAV vectors with optimal purity, potency, and consistency are needed. Some of the available rAVV purification methods include cesium chloride gradient ultracentrifugation, iodixanol gradient ultracentrifugation 3, heparin-based affinity chromatography purification(4) and ion-exchange chromatography 3. However, none of the above mentioned methods could solved one of the key drawbacks of the scalable rAAV purification process: contamination with empty capsids.
This work reports an optimized, scalable and reproducible method based on anion exchange chromatography to obtain high titers of rAAV vectors without empty capsids contamination.
MATERIALS AND METHODS
rAAV production and virus harvest
rAAV were obtained with AAV-DJ Helper Free Expression Systems from Cell Biolabs (USA). This system allows the obtaining of rAAV without the use of helper virus. For recombinant virus production, pAAV-DJ and pHelper were used as helper vectors and pAAV-GFP as genome vector. The method to produce rAAV-GFP was done following manufacturer's instructions. Briefly, 100-mm-diameter culture dishes were seeded with HEK-AAV cells at 70 % confluence, which were co-transfected with pAAV-GFP, pAAV-DJ and pHelper (1:1:1) with 25 kDa branched polyethylenimine.
Three days after transfection the cells were harvested by scrapping and were lysed with Tris-Cl buffer containing 150 mM NaCl and 2mM MgCl2. Virus-containing supernatant were precipitated adding 1/5 volume of PEG 40%/2.5M NaCl, incubated 1 hour in ice and spun down at 3000 g for 30 min. Both virus fractions were joined and subjected to three freeze-thaw cycles and treated with benzonaze nuclease and RNAse. The final spin down was done at 3000 g during 30 min and the virus-containing supernatant were collected.
Virus titration
The vector genome copies number (VG) was determined by qPCR using Brilliant III Ultra-Fast SYBRgreen qPCR Master Mix (Agilent). The viral DNA was extracted from 1, 10 and 20 μl of purified virus and was treated with 0.5 U DNase I (NEB) to digest any contaminating unpackaged DNA, followed by an additional treatmentwith 10 μg of proteinase K (Thermofisher Scientific) to assist in breaking capsids and releasing viral DNA. qPCR was run in an AriaMX (Agilent technologies) with primers annealing at AAV ITRs: forward primer 5´-GGA ACC CCT AGT GATGGA GTT-3´ and reverse primer 5´-CGG CCT CAG TGA GCG A-3. The program used was: 95ºC 10 min, then cycled 40 times at 95ºC for 15 seg, 60ºC for 30 seg and 72ºC for 30 seg. To generate the standard curve, a pAAV-GFP plasmid was used in serial dilutions from 1 × 109 to 1 × 102 genome copies (vg) and performed in triplicate. A no-template negative control was also performed in triplicate 1.
Anion Exchange Chromatography
Thirty milliliters of Q Sepharose Fast Flow (GE Healthcare) were loaded on XK16 column (GE Healthcare) and equilibrated with 5 column volume (CV) of equilibrium buffer (EB) (Tris-HCl 20 mM, NH4Ac 100mM). Eight milliliters containing 8x1012 vg were subjected to buffer interchange through Sephadex G25 molecular exclusion column and adjusted to 30 mL of final volume with EB. Virus sample was loading into the Q-Sepharose column at 8 mL/min. Virus sample was recirculated 3 times and then washed with 5 CV of EB.
In order to discard empty capsids and elute AAV particles, 3 different elution protocols were tested.
i: 120 mM to 500 mM NH4Ac linear gradient (5CV), 1 M NaCl (5CV).
ii: 150 mM NH4Ac (5CV), 500 mM to 1 M NH4Ac linear gradient (5CV), 1 M NaCl (5CV).
iii: 150 mM NH4Ac (5CV), 1M NaCl (5CV).
The elution profiles were checked for UV absorbance at 280 nm and all peaks were collected for further analysis.
SDS-PAGE
The electrophoresis was performed according to Laemmli 5 using a 12 % resolving polyacrylamide gel under reducing conditions with Coomassie blue staining. AccuRuler RGB Plus Prestained Protein Ladder was obtained from MaestroGen Inc.
rAAV/Empty capsid ratio determination.
In order to determinate rAAV/Empty capsid ratio, elution peaks from Q-sepharose chromatography were subjected to 260/280 nm in a SPECTROstarNano from BMG LABTECH. Before absorbance measurement, samples were treated with SDS 20% and heated at 75°C for 10 min 6.
RESULTS
Due to its well-known advantageous features in scalable production processes and proven capacity to bind AAV (6), the Q-Sepharose fast flow was chosen for the rAAV purification process.
Peaks 1 and 2 were obtained from the NH4Ac 120 to 500 mM linear gradient (fig.1). The 260/280-absorbance ratio showed that peak 1 corresponds to the empty capsids-containing fraction (Table1). Little increment in 260/280-ratio of peak 2 suggest that this 120 to 500 mM linear gradient can not resolve empty capsids and rAAV. On the other hand, NaCl 1M elution step resulted in a high titer fraction of rAAV.
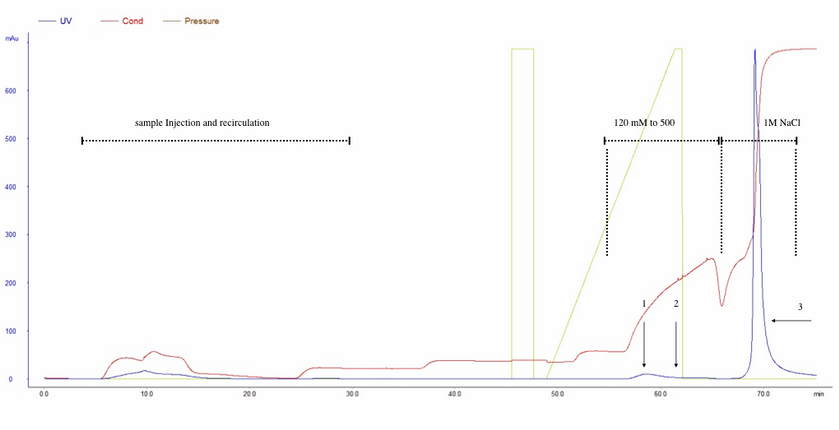
Figure 1. rAAV purification by anion chromatography interchange (Q-sepharose). Elution protocol i. 8x1012 vg were loaded into Q-sepharose XK-16 column and recirculated 3 times (8 mL/min). Empty capsids (1) and rAAV(2) were eluted by addition of a 120 to 500 mM NH4Ac linear gradient. 3: rAAV eluted with NaCl 1M solution.
Since empty capsid-containing fraction start to elute near to NH4Ac 120 mM, a wash step could be a suitable condition prior to rAAV elution. Taking into account previous findings, a 500 mM to 1 M NH4Ac linear gradient was tested followed by a NH4Ac 150mM wash step, in order to find out the best NH4Ac condition to elute rAAV. Finally, a high salt elution step (NaCl 1M) was tested, similar to the elution protocol i.
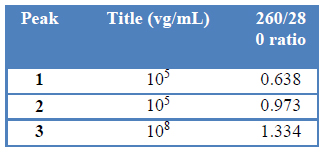
Table 1: Fraction analysis of rAAV Q-sepharose purification with elution protocol i.
A 150 mM NH4Ac wash step produced a first peak (peak1) (fig. 2) corresponding to the empty capsid-containing fraction. A 260/280 nm ratio analysis of this fraction confirmed that this condition is appropriated to wash out empty capsids (Table 2).
The application of a 500 mM to 1 M NH4Ac linear gradient generated three peaks at different NH4Ac molarities (fig2) and with different viral titers (Table 2). Once more, the fraction corresponding to NaCl 1M elution step (peak 5) resulted in a fraction with high rAAV titer.
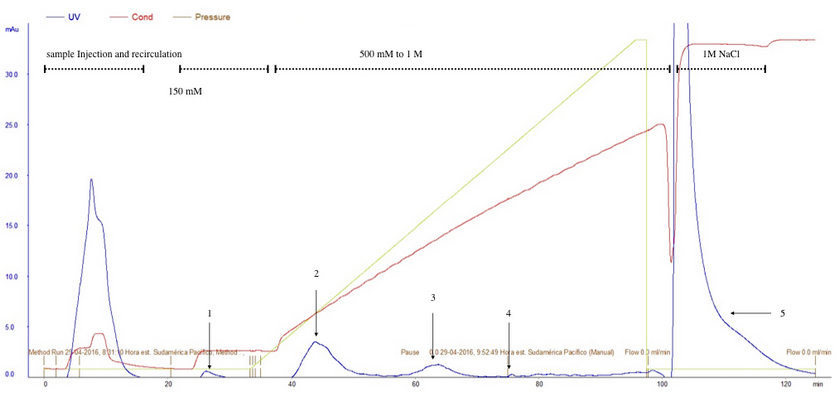
Figure 2. rAAV purification by anion chromatography interchange (Q-sepharose). Elution protocol ii. 8x1012 vg were loaded into Q-sepharose XK-16 column and recirculated 3 times (8 mL/min). 1: empty capsids were eluted by addition of 5 CV of NH4Ac 150 mM solution. 2,3,4: rAAV were eluted adding a 500 mM to 1M NH4Ac linear gradient. 5: rAAV eluted adding NaCl 1M solution.
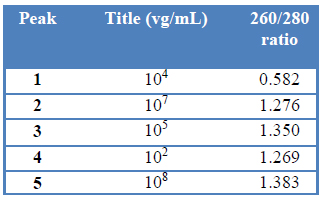
Table 2: Fraction analysis of rAAV Q-sepharose purification with elution protocol ii.
The 500 mM to 1 M NH4Ac linear gradient did not result in a homogeneous high viral titer fraction. Moreover, the high salt step condition generated a high titer rAAV fraction with elevated 260/280 ratio. Hence, a single high salt elution step could be sufficient to obtain a homogeneous rAAV fraction with high virus titer.
Figure 3 shows a chromatogram of the rAAV purification process, using two purification steps: a 150 mM NH4Ac wash step to eliminate empty capsids and a single high salt elution step with NaCl 1M.
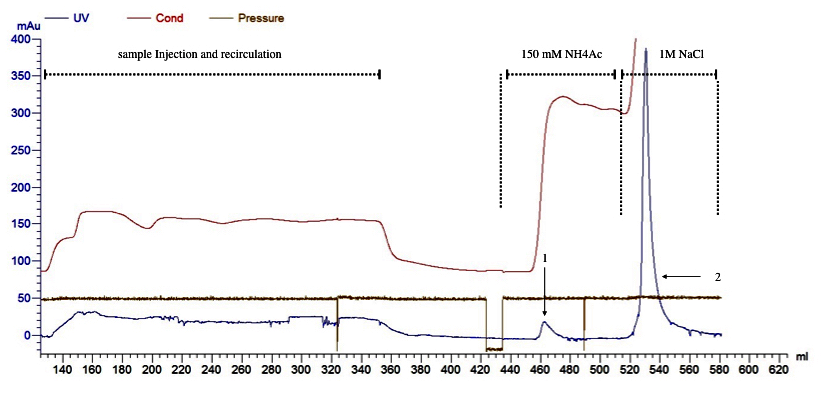
Figure 3. rAAV purification by anion chromatography interchange (Q-sepharose). Elution protocol iii. 8x1012 vg were loaded into Q-sepharose XK-16 column and recirculated 3 times (8 mL/min). 1: empty capsids were eluted by addition of 5 CV of NH4Ac 150 mM solution. 2: rAAV were eluted adding 5CV of NaCl 1M solution.
Like the previous elution protocols, NH4Ac 150 mM wash step resulted in a single peak (peak 1) enriched with empty capsids (Table 3). The fraction eluted with NaCl 1M was enriched with a high titer of rAAV. The SDS-PAGE analysis confirmed a rAAV-purified sample compared with the no treated sample obtained from the virus harvest step.
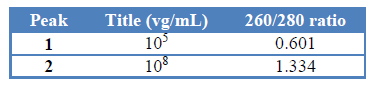
Table 3: Fraction analysis of rAAV Q-sepharose purification with elution protocol iii.
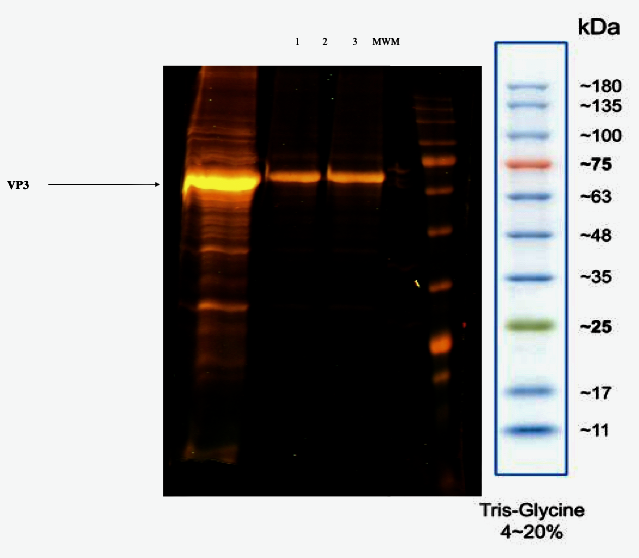
Figure 4. rAAV Q-sepharose purification. 12 % SDS-PAGE of eluted fractions with elution protocol iii. Lane 1: Injected rAAV sample. Lane 2: NH4Ac 150 mM wash step. Lane 3: NaCl 1 M elution step. MWM: AccuRuler RGB Plus Prestained Protein Ladder
DISCUSSION
Methods like Cesium chloride gradient ultracentrifugation and iodixanol gradient ultracentrifugation can resolve empty capsids and rAAV 3. However, these processes are time consuming, require expensive equipment and are not based on a scalable methodology. On the other hand, affinity based chromatography procedures like heparin-based affinity chromatography cannot solve contamination with empty capsids 4. Some authors have reported the use of both, anion and cation exchange chromatography to solve this problem 7, 8.
This work describes the development of a scalable purification method to remove empty capsids from hybrid capsid rAAVs. These hybrid rAAVs have a wilder tropism than other non-hybrids rAAVs.
Guang Qu et al. reported that the use of a NH4Ac 100 to 200 mM linear gradient as wash step and NH4Ac 200 to 300 mM linear gradient to rAAV2 elution, in a Poros HP50 column, resulted in a high yield of rAAV2 7. Similar results were also obtained with the application of a NH4Ac 180 mM in a Q-sepharose column 8. Although similar conditions were tested in this work, we could not be able to obtained similar results. Since hybrid rAAV carry eight different wild-type capsid viruses, the ionic strength could vary. Indeed, the application of NH4Ac linear gradient did not generate a homogeneous rAAV preparation.
In conclusion, a scalable method has been developed that allow the separation of empty capsids from hybrid rAAV with good purity yield. This method can be applied to other AVV serotypes.
Acknowledgment: We want to thanks to Fondecyt Grant # 3140153 for funding this work.
REFERENCES
1. Huang X, Hartley AV, Yin Y, Herskowitz JH, Lah JJ, Ressler KJ. AAV2 production with optimized N/P ratio and PEI-mediated transfection results in low toxicity and high titer for in vitro and in vivo applications. J Virol Methods. 2013;193(2):270-7.
2. Mietzsch M, Grasse S, Zurawski C, Weger S, Bennett A, Agbandje-McKenna M, et al. OneBac: platform for scalable and high-titer production of adeno-associated virus serotype 1-12 vectors for gene therapy. Hum Gene Ther. 2014;25(3):212-22.
3. Strobel B, Miller FD, Rist W, Lamla T. Comparative Analysis of Cesium Chloride- and Iodixanol-Based Purification of Recombinant Adeno-Associated Viral Vectors for Preclinical Applications. Hum Gene Ther Methods. 2015;26(4):147-57.
4. Clark KR, Liu X, McGrath JP, Johnson PR. Highly purified recombinant adeno-associated virus vectors are biologically active and free of detectable helper and wild-type viruses. Hum Gene Ther. 1999;10(6):1031-9.
5. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227(5259):680-5.
6. Sommer JM, Smith PH, Parthasarathy S, Isaacs J, Vijay S, Kieran J, et al. Quantification of adeno-associated virus particles and empty capsids by optical density measurement. Mol Ther. 2003;7(1):122-8.
7. Qu G, Bahr-Davidson J, Prado J, Tai A, Cataniag F, McDonnell J, et al. Separation of adeno-associated virus type 2 empty particles from genome containing vectors by anion-exchange column chromatography. J Virol Methods. 2007;140(1-2):183-92.
8. Guo P, El-Gohary Y, Prasadan K, Shiota C, Xiao X, Wiersch J, et al. Rapid and simplified purification of recombinant adeno-associated virus. J Virol Methods. 2012;183(2):139-46.
Received: 10 october 2018
Aproved: 2 december 2018
Camacho, F1., Cerro, R.P2., Varas, N1., Leiva, M.J2., Toledo, J.R2. and Sánchez. O1.
1 Pharmacology Department , Biological Sciences School, Universidad de Concepción.
2 Pathophysiology Department, Biological Sciences School, Universidad de Concepción.