2024..01.01.4
Files > Conference Series > 2024 > Chimboazo ild pagina nueva
Biological nematicides as an alternative for control of Meloidogyne incognita populations in yellow pitahaya (Sselenicereus megalanthus).

Yadira Vargas1*, Jimmy Pico1, Neiver Manobanda2, Angel Garcia1, Jessica Sanmiguel1.
1 Estación Experimental Central de la Amazonía, Instituto Nacional de Investigaciones Agropecuarias, La Joya de los Sachas, Ecuador; jimmy.pico@iniap.gob.ec; angel.garcia@iniap.gob.ec..; jessica.sanmiguel@iniap.gob.ec.
2 Tesista, Escuela Superior Politécnica de Chimborazo, Extensión Norte Amazónica;[email protected]
* Correspondence: yadira.vargas@iniap.gob.ec; +593 0991900401.
Available from. http://dx.doi.org/10.21931/BJ/2024.01.01.4
ABSTRACT
Yellow pitahaya in the Ecuadorian Amazon has become one of the most important economic crops in the region. However, the presence of pests (nematodes) in the soil has caused up to 100% of the crop's growth stages to be affected. Faced with this problem, growers use various chemical nematicides that minimize this impact but cause contamination problems. For this reason, the objective of the research was to evaluate microorganisms that control or reduce the population of Meloidogyne incognita in the pitahaya crop at the greenhouse level. The design was DBCA, and the statistical analysis was performed with the statistical package Infostat 2017, using linear mixed models and Fisher's tests at 5%. The results show that root nodules decrease when P. lilacinum + T. asperellum is applied after nematode injection (261). In addition, the lowest number of nodulations (251) was obtained when microorganisms were applied after nematode inoculation (251 nodulations). Still, when microorganisms were used before, aerial biomass growth was stimulated (384.17 g) even when nematodes were present in the root system.
Keywords: microorganisms; nematodes; pitahaya.
INTRODUCTION
In Ecuador, it is estimated that there are around 2000 hectares dedicated to pitahaya cultivation, mainly in the provinces of Pichincha, Manabí, and in the Amazon region in Morona Santiago, Orellana, and Sucumbíos 3. In the Amazon region, pitahaya cultivation is commercially a monoculture using conventional agronomic methods. This is mainly due to the vulnerability of pitahaya to various pests and diseases, such as fungi, bacteria, viruses, insects, and nematodes, which affect the plantations at all stages of growth. The most common nematodes affecting pitahaya belong to the genera Meloidogyne sp. (50-81%), Helicotylenchus dihystera (82-100%), Hemicycliophora sp., Tylenchorhynchus sp., Xiphinema sp., Trichodorus sp., Hoplotylus sp., Hemycicliophora sp., Dorylaimus (27%), Tylenchus (23%), Aphelenchus (14%), and Pratylenchus (5%) 4-5. In the Palora canton, which hosts the largest cultivated area of pitahaya, it has been observed that 97% of the plantations are affected by Meloidogyne sp. and Helicotylenchus spp.. In comparison, 3% are impacted by Tylenchus ssp. 6. Nematode infestation, especially of the genus Meloidogyne sp., in the root system of pitahaya plants causes a decrease in crop yield. This is due to the formation of nodules in the roots, which hinders water absorption and nutrients from the soil 7-8. Additionally, visible symptoms in the aboveground part of the plants include yellowing, thin and weak stems 9,6,5.
Currently, non-fumigant products, such as organophosphates and carbamates, chemical compounds with nematicidal activity, are used for nematode control in pitahaya cultivation. However, these products present environmental risks and can be toxic to humans 10. Studies conducted in different crops, such as lettuce, tomato, cauliflower, celery, and broccoli, have found pesticide residues (organophosphates and carbamates) in 48% of the analyzed products 11-13. Furthermore, it has been observed that 3% of agricultural workers exposed to pesticides suffer annually from chronic intoxication, neurological disorders, peripheral neuritis, male hormonal alterations, optic nerve problems, cataract formation, and respiratory effects 12.
Given that nematode control in pitahaya crops is mainly carried out with highly toxic nematicides, it is urgent to seek alternatives to the use of pesticides. In this regard, biological control through antagonistic organisms has been the research subject in recent years, and its potential for managing plant-parasitic nematodes has been recognized 14. These beneficial organisms include Pasteuria penetrans, Pasteuria hartismeri, Pochonia chlamydosporia, Bacillus firmus, Paecillomyces lilacinus, and Trichoderma spp. These microorganisms act by adhering to the nematodes' cuticle or parasitizing the females' eggs, resulting in the death of the nematodes. Although variable results have been obtained in research on antagonistic organisms, their effectiveness in controlling plant-parasitic nematodes has been demonstrated. However, only a tiny group of antagonistic organisms has been studied in detail 15.
For this reason, this research aimed to evaluate microorganisms that control or reduce the population of Meloidogyne incognita in the pitahaya crop at the greenhouse level. For this study, strains of microorganisms from pitahaya plantations, which have been selected at the in vitro level, were used. In addition, a commercial product based on P. lilacinum + T. asperellum was employed because there are products that no longer have viable fungal spores when they do not receive a good storage process.
MATERIALS AND METHODS
Location of the Experiment
This study was conducted at the Central Experimental Station of the Amazon (EECA), National Institute of Agricultural Research (INIAP), in Orellana, La Joya de los Sachas canton province. The experimental site was situated at 0291649 latitude and 09962311 longitude, with an altitude of 282 meters above sea level. The climate in the area is warm and humid tropical, with an average annual temperature of 25°C, an average maximum temperature of 22°C, an average minimum temperature of 40°C, and an average relative humidity of 90%. In the greenhouse, the average relative humidity is 70%, and the average temperature is 35°C.
Treatments
The experiment was organized in a randomized complete block design with three replications. The experimental unit consisted of eight pots with yellow pitahaya cuttings. The treatments consisted of T1 (Purpureocillium lilacinum) Laboratory strains, T2 (Trichoderma asperellum) Laboratory strains, T3 (Purpureocillium lilacinum + Trichoderma asperellum) Laboratory strains, T4 (Purpureocillium lilacinum + Trichoderma asperellum) Comercial product, T5 (absolute control), and T6 (control + nematode). The microorganisms stored in the laboratory and used for this study already come from a previous research process; they arrive from pitahaya plantations.
Specific Management of the Experiment
The study was implemented under greenhouse conditions. Yellow pitahaya cuttings of 40 cm were planted in pots with 4500 g of sterilized soil at 2 to 3 cm depth. After 22 days, the cuttings began to emit their first roots 16.
M. incognita was extracted from the galled roots of yellow pitahaya. The roots were washed, cut into approximately 1 cm sections, and blended with 100 ml of water in a blender for 20 seconds in two intervals with five seconds of rest. Subsequently, the blender content was passed through a set of nested sieves with 250, 150, and 25 µm openings (mesh sizes of 60, 100, and 500, respectively). The content placed on the 250 and 150 µm sieve was rinsed with running water for 1 minute. The sediment on the No. 500 sieve was collected in a graduated cylinder and filled with 100 ml of water. It was then homogenized with an air pump, and a 4 ml aliquot was taken for nematode identification and counting using a trinocular inverted microscope with LWD IOS objectives, X-LED illumination, and EWF10X/22mm eyepieces 17.
Finally, M. incognita was inoculated into 15-day-old tomato plants (Lycopersicum esculentum Mill) from transplant. After 30 days, nematodes were extracted from the L. esculentum Mill plants and inoculated into pitahaya plants. Approximately 1200 J2 nematodes were applied. Four 5 cm deep holes were made in the soil near the base of the plant stem for injection, the nematode solution was poured, and then the holes were covered 18,19.
The biological control agents used were T. asperellum and P. lilacinum. Spore crystals were taken with sterile forceps and seeded on Petri dishes containing Potato Dextrose Agar (PDA) medium 20. Sterilized rice substrates were used to mass produce the control agents and a conidial suspension of T. asperellum and P. lilacinum. The conidial suspension was prepared by adding 20 ml of sterile distilled water with 0.1% Tween 80 to each Petri dish containing T. asperellum and P. lilacinum with 6 days of growth. To this conidial suspension, 125 ppm of chloramphenicol was added. Subsequently, 3 ml of each suspension was taken using a sterile pipette and deposited in bags containing 150 g of the sterilized substrate (rice), which were then incubated for 15 days at a room temperature of 24 ± 2 °C 21. To establish the inoculum concentration, 1 g of rice from each multiplied substrate was weighed, and a suspension was prepared in 10 ml of distilled water. The spore quantification was performed by performing 20 readings in a Neubauer chamber 22. Once the concentration was determined, 1x10-9 spores were applied in 100 ml solutions per plant 7 days before and 7 days after the injection of M. incognita.
Evaluation Methods
At 30, 60, and 90 days after injection of M. incognita, a destructive evaluation was performed on 2 plants (treatment and replication). These plants were randomly selected, and the following parameters were assessed: incidence, severity, aboveground biomass weight, and final nematode population (PF).

Figure 1. Nematode Sampling in S. megalanthus Plants. a) Extraction of the plant and sampling .b) Register agronomic variables (plant and roots).
The number of plants infected by M. incognita was recorded 30, 60 and 90 days after inoculation with the nematode to determine the incidence. The results were expressed as a percentage (%) 23. To select the galling index (GI) in the root system of pitahaya plants, the number of galls formed in the root was counted and with the scale proposed by Taylor and Sasser 28 (0 to 6, where 0 = 0 galls; 1 = 1-2 galls; 2 = 3-10 galls; 3 = 11-30 galls; 4 = 31-100 galls; 5 = >100 galls) the severity was estimated 24.
All cladodes were collected per plant to determine aerial fresh weight (AFW). An analytical balance was initially used to define the fresh weight in grams (g). These procedures were carried out according to the study conducted by Gelpud et al. 25. The multiplication rate of nematodes in soil and roots (MR) was also determined by dividing the final population (Pf) by the initial population (Pi) Berroterán et al. 26.
Statistical analysis was performed with the statistical package Infostat version 2017, and analysis of variance was conducted using Generalized Linear Mixed Models. The difference between the means of the treatments was estimated using the Least Significance Differences (LSD) Fisher with a significance level of 5% 27.
RESULTS
For the nematode incidence variable, it was determined that all plants in the different treatments were affected by nematode infestation (ranging from 89% to 100%) (Table 1), demonstrating that the applied inoculum concentration (1200 J2) caused an infection process. The presence of nematodes in the absolute control may be attributed to potential contamination of the experimental units during irrigation.
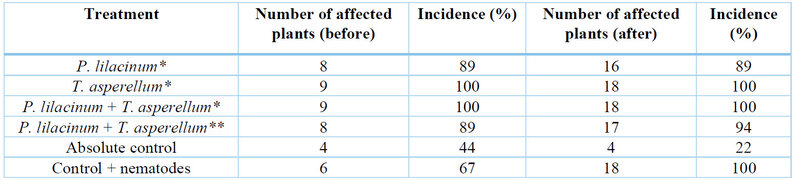
* Laboratory strains; ** Comercial product.
Table 1. Incidence of M. incognita in H. megalanthus plants.
A univariate analysis was performed for the variable "number of nodules" (Table 2). A highly significant difference was found for treatments (p<0.0001), a significant difference for the time of application "before and after," and the interaction (days*treatment) (p=0.0026; p=0.0170 respectively).
The main effect for treatments indicated that the lowest number of galls was formed when T. asperellum was applied. However, it was observed that the combination of the commercial product P. lilacinum + T. asperellum (1x 10-11 cfu/g) resulted in a lower number of galls compared to the control + nematode. The treatments where laboratory-obtained strains were applied (P. lilacinum + T. asperellum and P. lilacinum) at concentrations of 1x10-9 spores yielded the highest number of galls (Table 3). According to the scale reported by Taylor and Sasser 28, the severity grades for the different treatments were 5, meaning that the number of galls in the experimental units was high (more than 100 galls) (Table 3).

** significant at p ≤ 0.01, * significant at p ≤ 0.05.
Table 2. The main effects and interaction effect for the number of galls on roots determined for each factor: Treatment and Time of application.
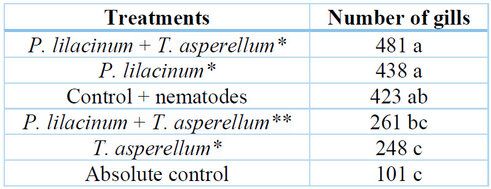
* Laboratory strains; ** Comercial product.
Table 3. Mean values of the number of galls in the root system of H. megalathus.
The analysis of the interaction treatment * season of application shows that the commercial product based on P. lilacinum + T. asperellum presents the least formation of nodulations at 60 and 90 days of evaluation. With T. asperellum the opposite happened, the lowest number of nodules was found at 30 days and as time passed the number of nodules increased (Table 4). On the other hand, the best time of application of the treatments was when the controllers were applied after the inoculation of the nematodes (251 nodulations) with respect to the 400 nodulations obtained when it was applied before (Table 4).
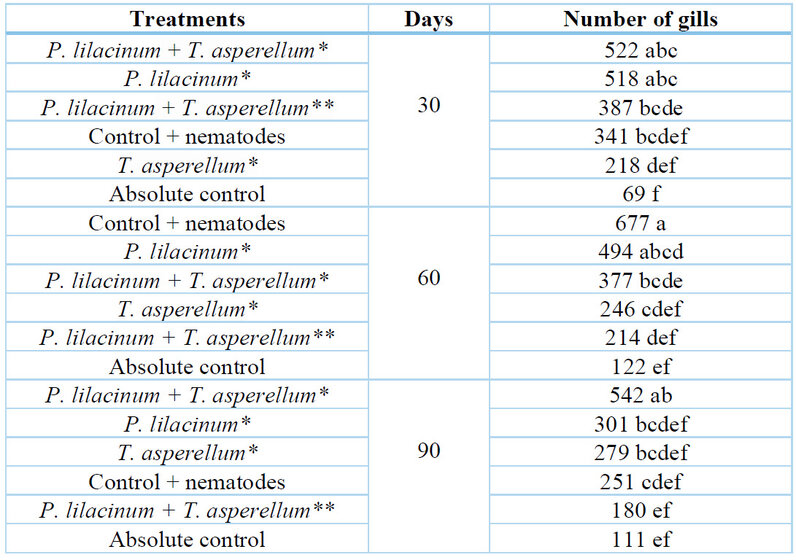
* Laboratory strains; ** Comercial product.
Table 4. Mean values of the number of galls by treatment and season of application.
The analysis in the air fresh weight (PFA) variable showed highly significant differences for the application time factor and treatments (p<0.0001) and not significant for the interaction (p=0.4632) (Table 5).

** significant at p ≤ 0.01; * significant at p ≤ 0.05; NS not significant.
Table 5. Main effects and interaction effect for aerial biomass determined for each factor: Treatment and Time of application.
It was determined that the amount of aerial biomass was higher when the biological controls were applied before the injection of nematodes (384.17 g) than when applied after (314.74 g). The main effect of the treatments showed that the amount of aerial biomass was higher when P. lilacinum + T. asperellum (obtained at laboratory level) was applied. On the other hand, it was observed that when T. asperellum obtained at the laboratory level was involved, the amount of aerial biomass was the lowest (Table 6).
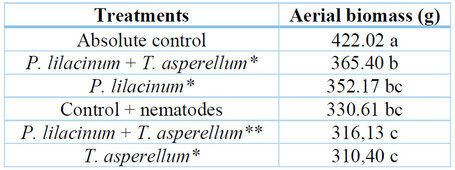
* Laboratory strains; ** Comercial product.
Table 6. Aboveground biomass (g) of H. megalanthus.
The analysis of the variable number of nematodes in the soil showed significant differences for the application times factor (p=0.0011) and non-significant differences for treatments and interaction (p=0.6139, p=0.2152, respectively). (Table 7).
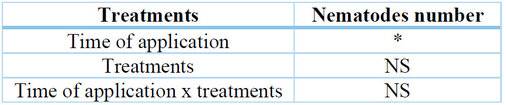
* significant at p ≤ 0.05, NS not significant.
Table 7. Main effects and interaction effect for the number of nematodes in soil determined for each factor.
Treatment and time of application.
The lowest amount of nematodes in the soil was found when the inoculation with the biocontrol agents was carried out before the inoculation of M. incognita (Table 8). However, this did not help to control the damage suffered by the roots of the pitahaya plants.
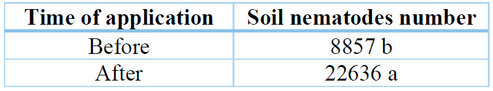
Table 8. Population of nematodes in the soil in two seasons of applications (before and after).
When analyzing the variable number of nematodes in the roots system of the pitahaya plants, significant differences were found for the time of application of the biocontrols (p=0.0198) and non-significant differences for the treatments and the interaction (p =0.5402, p=0.1429, respectively) (Table 9). That is, the biological controls, when applied before, did positively influence the population density of M. incognita (Table 10).
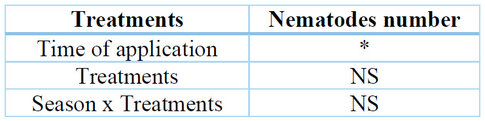
* significant at p ≤ 0.05, NS not significant.
Table 9. The main effects and interaction effect for the number of nematodes in S. megalanthus root were determined for each factor: Treatment and time of application.
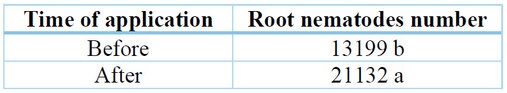
Table 10. Population of nematodes in the root system of S. megalanthus plants.
DISCUSSION
The number of galls in the root system of yellow pitahaya (S. megalanthus) indicates the plant's susceptibility to attack by M. incognita. In this study, the number of galls decreased by 62% and 70% when T. asperellum (laboratory strains) and P. lilacinum + T. asperellum (commercial) were applied, respectively. The excellent performance of Asperellum may be because it was collected in pitahaya plantations. It was also determined that applying P. lilacinum and P. lilacinum + T. asperellum strains obtained in the laboratory produced 4% and 13% more galls than the control + nematodes. This behavior could be attributed to certain micro-organisms possessing root growth-promoting properties, especially micro-organisms of the genus Trichoderma. This behavior could potentially be attributed to the root growth-promoting properties of the Trichoderma genus. This behavior is supported by Brotman et al. 29, who mentioned that Trichoderma activates auxins and several genes responsible for plant root development. However, Kariga et al. 30 mention that when placed, T. asperellum M2RT4 reduced the number of galls, egg mass, and nematode eggs. On the other hand, when both microorganisms are applied together, they seem to exert more efficient control, as T. asperellum T203 colonizes the roots and P. lilacinum reduces root nodulation, mass and egg generation while favoring host growth. 30.
At 30 days, the presence of T. asperellum minimized nematode attack on the pitahaya root system, possibly because the concentration decreased by 26% at 60 days 31. At 60 and 90 days, it was observed that P. lilacinum + T. asperellum (commercial) resulted in lower nodule formation compared to the control + nematode and the laboratory-obtained strains of T. asperellum and P. lilacinum. This may be attributed to the stable germination of the microorganisms 31. The lowest amount of galling was observed when the organisms were applied after nematode inoculation, which differed from the reported behavior of reducing root galling, egg mass, and egg production when the microorganisms were applied ten days before (reference not provided).
Applying the microorganisms seven days earlier positively influenced plant growth (aboveground biomass). This is possible because the microorganisms stimulate the growth of the root system. Kariga et al. 30 noted that the application of T. asperellum M2RT4 and P. lilacinum (MR2 and KLF2) increased host growth and reduced nematode populations in soil and roots. Silva 32 also determined that P. lilacinus and several Trichoderma species promoted root development, growth and plant production when the microorganisms were applied before Meloidogyne inoculation. Also, it was resolved that plants had higher aerial biomass when inoculated with P. lilacinum + T. asperellum and laboratory-derived T. asperellum. This behavior may be attributed to these microorganisms being collected in pitahaya plantations in the Ecuadorian Amazon, which is why they performed better. Rodriguez-Kabana et al. 33 highlight the substantial divergence in the behavior of microorganisms, particularly concerning their efficacy in biocontrol and their proficiency in establishing themselves within the soil. This underscores the necessity of ensuring harmonious compatibility with distinct local conditions. For instance, Ortiz et al. 34 noted an illustrative case wherein the application of P. lilacinum on Psidium guajava to manage Meloidogyne spp. did not exhibit any detrimental impact on plant growth and development, even when nematodes were present within the root system. This finding is compelling in demonstrating that the behavior of microorganisms differs especially if they are used in other environments and are not native.
CONCLUSIONS
The application of the commercial dosage of P. lilacinum + T. asperellum led to a decrease in the number of galls present on the roots as compared to the untreated control. Nonetheless, no notable enhancement in plant growth was noted. Conversely, plant development was increased when employing a dosage of the laboratory-derived P. lilacinum + T. asperellum strain; however, the gall count did not decrease. These findings indicate that the strains under examination in this study possess constrained potential for nematode control.
This study established the importance of searching for new local strains and adjusting application rates since the behavior of microorganisms, especially in biocontrol and establishment capacity, can vary significantly from one place to another.
Author Contributions: Conceptualization, Y.V.-T., JP; methodology, Y.V.-T. and JP; statistical analysis, Y.V.-T., JP, writing—original draft preparation, Y.V.-T.; writing—review and editing, Y.V.-T., JP, NM, JS and AG All authors have read and agreed to the published version of the manuscript.
Funding:
This research was financed by the National Institute of Agricultural Research (INIAP) through the Central Experimental Station of the Amazon (EECA) and by the Research Fund for Agrobiodiversity, Seeds and Sustainable Agriculture (FIASA).
Institutional Review Board Statement:
Does not apply.
Informed Consent Statement:
Does not apply.
Data Availability Statement:
The data is contained in the article.
Acknowledgments:
This work was completed with the support of agronomists from the Fruit Programme of the Central Experimental Station of the Amazon.
Conflicts of Interest:
The authors declare no conflict of interest.
REFERENCES
1. Verona-Ruiz A, Urcia-Cerna J, Paucar-Menacho LM, Verona-Ruiz A, Urcia-Cerna J, Paucar-Menacho LM. Pitahaya (Hylocereus spp.): Cultivo, características fisicoquímicas, composición nutricional y compuestos bioactivos. Sci Agropecu 2020, 11(3):439-53.
2. Zúñiga Rizo CA. Biología floral y estructura vegetativa de la pitahaya (Hylocereus undatus Haworth) 2016.
3. Vargas Tierras YB, Pico JT, Díaz M. A, Sotomayor Akopyan DA, Burbano A, Caicedo V. C, et al. Manual del Cultivo de Pitahaya para la Amazonía Ecuatoriana 2020.
4. Castaño SP, Rincón A, Varón de Agudelo F. Identificación de nematodos asociados con pitaya Acanthocereus pitahaya Jaq. Dugand. Rev ICA 1991, Vol 26 Núm 1-2.
5. Guzmán-Piedrahita ÓA, Pérez L, Patiño A. reconocimiento de nematodos fitoparásitos en pitahaya amarilla (Selenicereus megalanthus HAW.). Bol Científico Cent Mus Mus Hist Nat 2012, 16(2):149-61.
6. Delgado A, Pico JT, Navia D, Suárez C. Memorias: Nemátodos fitoparásitos asociados al sistema radical del cultivo de pitahaya amarilla en el cantón Palora. EC: INIAP, Estación Experimental Central de la Amazonía 2019.
7. Cedeño Ronquillo AI. Ocurrencia de nematodos fitoparásitos en el cultivo de Pitahaya Selenicereus megalanthus en la zona de Las Naves, provincia de Bolívar. Facultad de Ciencias Agrarias Universidad de Guayaquil; 2021.
8. Osorno YCR, Hoyos LFP, Sánchez DAC. Efecto de los nematodos en la cantidad y calidad de raíces y métodos de evaluación. Rev Politécnica 2008.
9. Haro Altamirano JP, Zambrano Cárdenas GO, Cazorla Vinueza XR, Soplín Villacorta H, Garzón R. Evaluación de la sustentabilidad de fincas de la agricultura familiar, de dos eco tipos de Pitahaya amarilla (Selenicereus megalanthus), y su subproducto. Polo Conoc Rev Científico - Prof 2021.
10. Obando Vergara M, García Morera G, Araya M. Control químico de Globodera pallida (Stone) Behrens y la producción de papa (Solanun tubersoum L.), variedad Floresta. Rev Protección Veg 2017, 32(3):00-00.
11. Aguilar Calizaya VS, Heredia Araujo J. Determinación de la concentración de agroquímicos en productos hortícolas en la Localidad de Carapongo-Lima-Perú-2020. Univ Peru Unión 2020.
12. Ordoñez Beltrán V, Frías Moreno MN, Parra Acosta H, Martínez Tapia ME. Estudio sobre el uso de plaguicidas y su posible relación con daños a la salud. Rev Toxicol 2019, 36(2):148-53.
13. Karel, Zabala-Velin AA, Villarroel-Quijano KL, Sarduy-Pereira LB. Evaluación del impacto ambiental del cultivo de la pitahaya, Cantón Palora, Ecuador. TecnoLógicas 2020, 23(49):92-107.
14. Varela-Benavides I, Durán-Mora J, Guzmán-Hernández T. Evaluación in vitro de diez cepas de hongos nematófagos para el control de Meloidogyne exigua, Meloidogyne incognita y Radopholus similis. Rev Tecnol En Marcha 2017, 30(1):27-37.
15. Bishop AH, Gowen SR, Pembroke B, Trotter JR. Morphological and molecular characteristics of a new species of Pasteuria parasitic on Meloidogyne ardenensis. J Invertebr Pathol 2007, 96(1):28-33.
16. Rahad M, Islam M, Rahim M, Monira S. Effects of rooting media and varieties on rooting performance of dragon fruit cuttings (Hylocereu sundatus Haw.). ResearchGate 2016, 3(1):67-77.
17. Gilces CT, Santillan DN, Velasco L. Plant-Parasitic Nematodes Associated with rice in Ecuador. Nematropica 2016, 46(1):45-53.
18. Hussey R, Janssen G. Root-knot nematodes: Meloidogyne species. Plant Resist Parasit Nematodes 2022, 43-70.
19. Palacino JH, Leguizamon C, Jairo. Interaction between Glomus manihotis and Melidogyne incognita in yellow and red pitaya, under nursery conditions. Fitopatol Colomb 1991, 15(1):9-17.
20. Hoyos Carvajal LM, Jaramillo Villegas S, Orduz Peralta S. Evaluación de Trichoderma asperellum como biorregulador de Spongospora subterranea f. sp. subterranea. Rev Fac Nac Agron Medellín 2008, 61(2):4496-502.
21. Alcantara YML. Producción y formulación de trichoderma asperellum para el manejo de patógenos de la raíz de caña de azúcar 2019.
22. Ramírez HG, Granja AZ, Aguila E, Cantoral MT. Manual de producción y uso de hongos entomopatógenos. Lab Entomopatógenos SCB-SENASA 2014.
23. Álvarez S DE, Botina J JA, Ortiz C. AJ, Botina J LL. Nematicide evaluation of the essential oil from Tagetes zypaquirensis in the control of the nematode Meloidogyne spp. Rev Cienc Agríc 2016, 33(1):22-33.
24. Chaves CG, Marcillo EM, González CS, Garcia CB. Susceptibilidad de genotipos de Solanum spp. al nematodo causante del nudo radical Meloidogyne spp. (chitwood). Acta Agronómica 2011, 60(1):50-67.
25. Gelpud C, Mora E, Salazar C, Betancourth C. Susceptibilidad de genotipos de Solanum spp. al nematodo causante del nudo radical Meloidogyne spp. (chitwood). Acta Agronómica 2011, 60(1):50-67.
26. Berroterán G, Perichi G, Aguirre Y. Reacción de seis genotipos de caraota (Phaseolus vulgaris L.) al nematodo agallador Meloidogyne javanica (Treub) Chitwood. Bioagro 2020, 32(1):67-71.
27. Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L, Tablada M, Robledo CW. Infostat versión 2018. 2015. Disponible en: http://www.infostat.com.ar/
28. Taylor A, Sasser J. Biología, identificación y control de los nematodos de nódulo de la raíz. Univ Carol Norte 1983;111.
29. Brotman Y, Landau U, Cuadros-Inostroza Á, Takayuki T, Fernie AR, Chet I, et al. Trichoderma-Plant Root Colonization: Escaping Early Plant Defense Responses and Activation of the Antioxidant Machinery for Saline Stress Tolerance. PLoS Pathog 2013, 9(3):e1003221.
30. Kiriga AW, Haukeland S, Kariuki GM, Coyne DL, Beek NV. Effect of Trichoderma spp. and Purpureocillium lilacinum on Meloidogyne javanica in commercial pineapple production in Kenya. Biol Control 2018.
31. Viera W, Noboa M, Bermeo J, Báez F, Jackson T, Viera W, et al. Parámetros de calidad de cuatro tipos de formulaciones a base de Trichoderma asperellum y Purpuricillium lilacinum. Enfoque UTE 2018, 9(4):145-53.
32. Silva Zegarra JA. Eficiencia de cinco especies de hongos nematófagos sobre el control de Meloidogyne spp. en pimiento del piquillo (Capsicum annuum L.) bajo condiciones de casa malla. Univ Priv Antenor Orrego 2021.
33. Rodriguez-Kabana R, Morgan-Jones G, Godoy G, Gintis BO. Effectiveness of species of Gliocladium, Paecilomyces and Verticillium for control of Meloidogyne arenaria in field soil. Nematropica. 1984, 14(2):155-70.
34. Ortiz Paz RA, Guzmán Piedrahita ÓA, Leguizamón Caycedo J. Manejo integrado del nematodo del nudo radical [Meloidogyne incognita (Kofoid & White) Chitwood Y Meloidogyne mayaguensis Rammh & Hirschmann] en almácigos de guayabo (Psidium guajava Linneo), variedad palmira ICA-1. Bol Científico Cent Mus Mus Hist Nat 2015, 19(2):104-38.
Received: 9 October 2023/ Accepted: 15 January 2024 / Published:15 February 2024
Citation. Vargas Y, Pico J, Manobanda N, Garcia A, Sanmiguel J.Biological nematicides as an alternative for control of Meloidogyne incognita populations in yellow pitahaya (Sselenicereus megalanthus). Revis Bionatura 2024; 1 (1) 4. http://dx.doi.org/10.21931/BJ/2024.01.01.4
Additional information Correspondence should be addressed to yadira.vargas@iniap.gob.ec
Peer review information. Bionatura thanks anonymous reviewer(s) for their contribution to the peer review of this work using https://reviewerlocator.webofscience.com/
All articles published by Bionatura Journal are made freely and permanently accessible online immediately upon publication, without subscription charges or registration barriers.
Publisher's Note: Bionatura Journal stays neutral concerning jurisdictional claims in published maps and institutional affiliations.
Copyright: © 2024 by the authors. They were submitted for possible open-access publication under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).